Глава 15: Триасовый период.
Триасовый период (триас) — первый геологический период мезозойской эры. Начался 251,902±0,024 млн. лет назад, закончился 201,3±0,2 млн. лет назад. Продолжался, таким образом, около 51 млн. лет. Название «триасовый период» происходит от греч. trias (троица) в связи с четким трехчленным строением толщи отложений этого периода в северной Германии. В основании толщи залегают красноцветные песчаники, в середине – известняки, а вверху – красноцветные песчаники и глинистые сланцы. Во время триаса биологическое разнообразие восстанавливается, возникают новые виды растений, рептилий и насекомых, формируется класс динозавров.
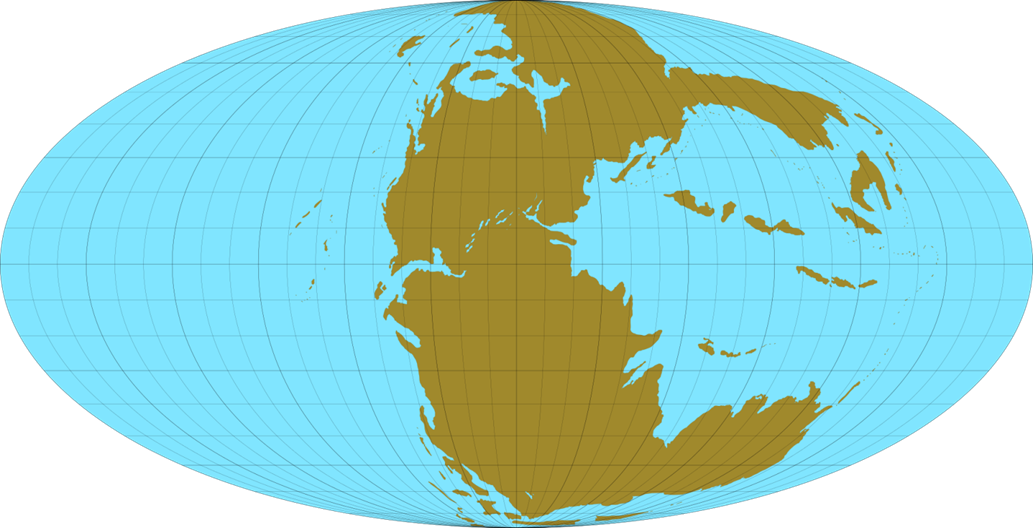
Климат и география.
До наступления триаса все материки составляли единый гигантский суперконтинент — Пангею. В конце триаса началось разделение Пангеи на два континента - Лавразию и Гондвану, которое завершилось в юрский период. Это сопровождалось бурной вулканической активностью, которое привело к поднятию общей материковой массы и пересыханию множества озер и рек. Во внутренних областях материков образовались обширные пустыни. Не смотря на это, на континенте еще оставалось много водных бассейнов. Что касается океанов, которые можно было наблюдать в триасовый период, их было только два:
- Панталасса: самый большой океан на Земле. Он окружил всю Пангею и был очень глубоким.
- Тетис: Он был расположен в восточной части Пангеи как своего рода залив, из которого в дальнейшем образовался Атлантический океан.
Климат, на большей части Пангеи, был сухой с очень жарким летом и холодной зимой и был в целом теплее, чем в наше время, без полярных ледяных шапок. Чем дальше от экватора, тем более умеренным он становился, а сезонный муссонный климат преобладал ближе к прибрежным регионам. В начале триасового периода концентрация кислорода в атмосфере составляла 10—15 %. Около 215 млн. лет назад за период около 3 млн. лет, что очень быстро в геологическом плане, уровень кислорода в атмосфере подскочил с 15 % до 19 %.
Животный мир.
После массового Пермского вымирания лишь немногие семейства животных перешли из палеозойской эры в мезозойскую. Первые 10 миллионов лет биосфера триасового периода мучительно восстанавливалась от катастрофы. Благодаря вымиранию освободилось множество экологических и эволюционных ниш, которые вновь стали бурно заполняться животным разнообразием. Ввиду того что в первой половине триасового периода климатические условия были почти одинаковыми на всей территории суши, животный мир был почти идентичен в своем разнообразии на всех участках суперконтинента. Сезонные изменения температур, во второй половине триасового периода, стали оказывать заметное влияние на растения и животных. Отдельные группы пресмыкающихся смогли приспособиться к холодным сезонам. Именно от этих групп произошли млекопитающие, а несколько позднее — и птицы.
Водная фауна.
Среди беспозвоночных триасового периода, в водах морей и океанов, уже имеются все типы животных, которые существуют и в наше время. Самые распространённые – это рифообразующие организмы. В палеозое уже существовали животные, которые колониями покрывали дно моря, образуя рифы, хотя и не очень мощные. В триасовый период, когда вместо табулят появляется много колониальных шестилучевых кораллов, начинается формирование рифов толщиной до тысячи метров. В результате массового развития и быстрого роста кораллов на дне моря образовывались подводные леса, в которых селились многочисленные представители других групп организмов. Двустворчатые моллюски, водоросли, морские ежи, морские звезды, губки жили между кораллами. Некоторые из них тоже принимали участие в рифообразовании.
Довольно характерны для триасового периода двустворчатые моллюски. Их тонкие, как бумага, раковины с хрупкими ребрами образуют в отдельных случаях целые прослойки в отложениях данного периода. Жили двустворчатые моллюски в мелководных илистых бухтах — лагунах, на рифах и между ними. В верхнетриасовом периоде появляется множество толстораковинных двустворчатых моллюсков, прочно прикреплявшихся к известняковым отложениям мелководных бассейнов.
Самыми распространенными из брюхоногих моллюсков были переднежаберные. По-прежнему доминируют головоногие, при этом доминирующий подкласс триасовых головоногих – аммониты. Некоторые из них достигали исполинских размеров с диаметром раковин до 5 м. Правда, и теперь в морях обитают исполинские головоногие моллюски, например кальмары, достигающие 18 м в длину, однако в мезозойскую эру исполинских форм было намного больше.
Аммониты, впервые появившиеся на Земле в девонский период, наиболее изученные и часто встречающиеся ископаемые морские животные. Они являются предшественниками современных осьминогов и кальмаров. Своего наивысшего расцвета, если иметь в виду их численность и разнообразие, аммониты достигли в пермский период. Затем в конце этого периода они почти полностью исчезли при массовом вымирании. Но разделаться с аммонитами оказалось не так-то просто. Некоторые из них сумели дожить до триасового периода и благодаря своим склонностям к дальним странствиям вскоре вновь распространились по всему миру. К середине мезозоя они достигли нового пика эволюционного процветания. И все же подобное процветание не могло быть вечным, и в конце мелового периода все аммониты внезапно исчезли с лица Земли — вместе со множеством других морских животных, включая белемнитов, плиозавров, ихтиозавров и плезиозавров.

Невероятное изобилие ископаемых останков аммонитов оказалось несколько обманчивым, заставив ученых в свое время сделать не совсем верные выводы об их распространении в прошлые эпохи. До самого последнего времени специалисты полагали, что аммониты населяли все первобытные моря без исключения. Однако затем было установлено, что отнюдь не все их разновидности обитали по всему земному шару. У разных видов были различные ареалы, что, по всей вероятности, было связано с рядом физических факторов, например с температурой воды или с процентным содержанием в ней соли (соленостью).
Смещение материков в конце мелового периода привело к грандиозным климатическим изменениям и общей трансформации (преобразованию) природной среды, затронувшей и Мировой океан. В морях появились новые моллюски, такие, как устрицы. Они зарывались в донный песок, пропускали через свои раковины воду и отфильтровывали из нее частицы пищи. Появилось и множество новых брюхоногих моллюсков (улиток и их родственников). С понижением уровня воды в мелких морях их скалистые берега все более обнажались. Здесь находили себе пристанище новые виды моллюсков, например блюдечки и литорины. Встречались и новые разновидности кораллов, креветок и омаров. Представителями иглокожих были морские ежи (предки современных ежей), лилии и морские звезды. В морях обитали губки, мшанки, листоногие раки, остракоды.
Появляются первые примитивные костистые рыбы. Мощные плавники, хорошо развитый зубной аппарат, совершенная форма, прочный и легкий скелет — все это способствовало быстрому распространению костистых рыб в морях нашей планеты. Дальше от берега в открытом море обитали наиболее высокоорганизованные виды рыб. Выжившие акулы и костные рыбы оспаривали друг у друга добычу. Со временем у них развились мощные челюсти способные разгрызать панцири крабов и раковины любых моллюсков.
Кистеперых рыб по-прежнему немного, в основном это целаканты и двоякодышащие. Резко прибавили в разнообразии лучеперые рыбы, наконец-то они смогли составить достойную конкуренцию другим классам рыб. Среди лучеперых появились хищники наподобие современной щуки, крупнейшие триасовые лучеперые рыбы достигали 1 м в длину. Заурихтис среди прочей пищи употреблял птерозавров, их окаменевшие останки находили в желудках этих рыб. Хрящевые рыбы в триасе сильно сдали позиции. Примитивные бесчелюстные конодонты, известные начиная с конца кембрия, в триасе окончательно вымирают.
В Триасовый период происходят активные изменения водной фауны. В раннем триасе вымирают криптостаматы и трепастоматы из мшанок, и число семейств этой группы становится минимальным за всю историю их существования. Значительное вымирание происходит среди головоногих моллюсков. В середине периода перестают существовать агониатиты, а к концу - цератиты из аммоноидей. К концу этого отрезка времени исчезают ортоцератиды и бактритоидеи, а также все палеозойские группы наутилид.
Одновременно с исчезновением шло и появление новых групп. Так, среди головоногих моллюсков еще в конце раннего триаса появляется отряд филлоцератид. Вымерло и появилось несколько отрядов рыб. В частности, исчезли последние брадиоданты из цельноголовых, появляются и вскоре исчезают три новых отряда в надотряде палеонисков, становятся известны два новых отряда среди хрящевых ганоидов и два отряда из костных ганоидов. В целом картина развития водной фауны в триасе довольно сложная и говорить о каком-либо единовременном сильном изменении в этой группе организмов не приходится.
Насекомые.
В триасе происходят изменения в классе насекомых. Например, сетчатокрылые виды в течение всего периода становились более разнообразными. В раннем триасе не было наземных жуков, хотя в перми они были широко распространены. В среднем триасе стали расцветать полужесткокрылые, жесткокрылые - стрекозы и подёнки. В позднем триасе появляется один из последних больших отрядов насекомых - двукрылые, прямокрылые, а также перепончатокрылые. Кроме того появились титаноптеры (titanoptera) – огромные кузнечики до 36 см в длину. В конце триаса вымерло одно из восьми семейств стрекоз.
В конце триасового периода климат стал жарким и засушливым. Мелкие чешуекрылые из-за увеличенного соотношения поверхности и объема тела особенно активно теряли жидкость во время полета. Поэтому у них появились хоботки для быстрого поглощения капелек росы, сока и других жидкостей. С их помощью они наверняка высасывали опылительные капли голосеменных, которые те выделяли для улавливания зерен пыльцы. Во всяком случае, современные хоботковые чешуекрылые посещают с этой целью гнетовидные растения. Особой пользы мезозойским голосеменным такие визиты не приносили, поскольку первые чешуекрылые не участвовали в опылении. И только с возникновением цветковых растений в середине мела чешуекрылые возложили на себя роль опылителей.
Амфибии.
Небольшую часть животных в триасе занимали амфибии или земноводные. Лепоспондилы вымерли в пермско-триасовом вымирании, остались только темноспондилы. Самая большая группа триасовых темноспондилов – капитозавры – крупные крокодилообразные существа от 2 до 6 м в длину. Земноводные также были представлены стегоцефалами из группы лабиринтодонтов. Это были малоподвижные животные с небольшим туловищем, маленькими конечностями и крупной головой. Они лежали в воде в ожидании добычи, и когда добыча приближалась, хватали ее. Их зубы имели сложную лабиринтообразную складчатую эмаль, поэтому их и назвали лабиринтодонтами. Кожа увлажнялась слизистыми железами. Наиболее характерные представители лабиринтодонтов — мастодонозавры. Эти животные, черепа которых достигали одного метра в длину. Они охотились на рыб и поэтому редко оставляли водную среду. Болот становилось меньше, и мастодонозавры вынуждены были заселять все более глубокие места, часто скапливаясь в большом количестве. Вот почему теперь находят много их скелетов на небольших участках.
Капитозавры
Среди амфибий также известны трематозавры – мелкие существа, до 60 см в длину и плагиозавры – забавные существа, сохранявшие во взрослом состоянии наружные жабры, как у головастиков. В длину они достигали 1 м. Также можно упомянуть метопозавров достигавших 3 м в длину и чигутизавров. В триасе появилась первая лягушка – триадобатрах. В отличие от современных лягушек, у этого 10-сантиметрового животного был маленький рудиментарный хвостик. Позже к амфибиям присоединились первые сухопутные и водные черепахи (истинные черепахи, а не плакодонты), пока с не полностью еще окостеневшим панцирем, под который пока еще нельзя прятать голову и лапы.
Рептилии.
Многие рептилии, обитавшие на Земле в пермский период, вымерли к началу триаса. Но некоторые из зверообразных рептилий все же уцелели. Количество растительноядных рептилий стало быстро увеличиваться. Некоторые популяции, такие, к примеру, как стадные листозавры, достигли небывалой численности. Их по праву называли бегемотами триасового периода, поскольку они вели образ жизни очень похожий на жизнь нынешних бегемотов. Листозавры также нежились на солнышке вокруг болот и прочих мелких континентальных водоемов, а в сильный зной забредали в воду, чтобы в ней слегка охладиться. Вот только численность данных стад не в пример нынешним бегемотам была просто громадна. Останки данных животных были найдены во всех уголках земного шара. Такое же увеличение численности произошло и с некоторыми хищниками. В скором времени это громадное количество животных переросло в качество и дало множество новых и необычных видов.
Листозавр
Одним из видов рептилий переживших пермскую катастрофу и живших в триасовый период были пеликозавры - звероподобные рептилии. Их название переводится как "Парусные ящеры". Спину пеликозавров украшал веерообразный гигантский парус. Одним из самых ярких и удивительных пеликозавров был диметродон.
На смену пеликозаврам вскоре пришла другая группа рептилий их называют терапсиды. Некоторые из них были растительноядными, а другие хищниками. Лапы терапсид были длиннее, чем у пеликозавров, а хвосты короче. Терапсиды были подвижнее и успешнее пеликозавров. В их физическом аспекте эти животные казались гибридом между собакой и ящерицей. Они были четвероногими, их конечности были немного длинными, а хвосты короткими. Первыми появились травоядные, а затем и плотоядные. Важность терапсидов заключается в том, что, согласно исследованиям, они были прямыми предками млекопитающих.
В среднем триасе появилось много удивительных рептилий. Например: представитель подкласса синаптозавров - трилофозавр лазил по деревьям и питался растительной пищей, а по внешнему виду напоминал кошку. Эупаркерииды (до 55 см вместе с хвостом) были похожи на мелких динозавров, но не родственны им. Это были очень проворные существа, есть мнение, что они могли перебегать водоемы по поверхности, как современные ящерицы-василиски.
Также в среднем триасе образовалась еще одно ответвление рептилий, по строению опорно-двигательной системы схожее с арзозаврами, но имеющее несколько иное строение челюстей. Назывались они рихозаврами и имели на концах челюстей массивные клювы, за что и были прозваны «клювокрылыми». Строение их челюстей, а также зубо-клювое их оснащение позволяло данным рептилиям не только кусать и жевать, но и преспокойно перерезать и перерубать добычу. Вдобавок ко всему при закрытии оконечный острый выступ нижней челюсти входил в специальный желоб верхней наподобие перочинного ножа, складывающегося в рукоять. При таком раскладе жертва была обречена.
Львиную долю всех описанных триасовых пресмыкающихся составляют рептилиоморфы. Самая большая группа триасовых рептилиоморфов принадлежит к подклассу диапсидов, это наиболее продвинутые пресмыкающиеся с облегченным черепом. По сравнению с пермским периодом их разнообразие увеличилось на порядок. Одной из групп триасовых диапсидов являлись лепидозавроморфы - предки всех современных ящериц и змей.
Наиболее процветающая группа диапсид – архозавроморфы, а самая процветающая ее подгруппа – архозавры то есть "правящие рептилии". Они появились в начале триасового периода и были предками всех основных групп известных в мезозойскую эру. Потомками архозавров были все виды динозавров, крокодиломорфы, нотозавры, плиозавры и плезиозавры, ихтиозавры, плакодонты и птерозавры.
Первые архозавры были маленькими зверьками, которые охотились за мелкой дичью по берегам озер и рек. Впоследствии из них развились куда более крупные животные. Эти существа имели особое строение бедра, что позволяло им легко вставать на задние ноги. В условиях, когда четвероногая походка была еще не отработана (горгонопсы вымерли, а больше никто нормально бегать не умел), передвижение на двух ногах было серьезным преимуществом, оно позволяло передвигаться заметно быстрее. Впрочем, в триасе большинство архозавров на двух ногах только бегали, а ходить предпочитали на четырех.
В первую очередь мы рассмотрим самую большую группу триасовых архозавров – круротарсов (crurotarsi). В отличие от первых динозавров, это были коренастые, мощные и относительно неповоротливые существа. Однако представители самого большого отряда круротарсов – райзухи внешне почти не отличались от хищных динозавров юрского периода. Самое большое семейство райзухов – райзухиды, сюда входят крупные хищники длиной 3-10 м. Другими похожими на райзухид рептилиоморфами, являлись попозавриды, они отличались от райзухид меньшими размерами (до 2 м) и большей склонностью к передвижению на двух ногах. Сюда также входили - ктенозаурисциды (до 2,6 м) отрастившие на спине парус для терморегуляции.
Ближе к концу триасового периода многие наземные животные, появившиеся в его начале, вымерли. Их место заняли новые рептилии, развивавшиеся на протяжении триаса. Примерно 225 млн. лет назад возникла группа рептилий, которых назвали текодонтами ("ячеистозубыми"). Поначалу это были неуклюжие приземистые животные, немного похожие на крокодилов. Они вели водный образ жизни и плавали при помощи мощного хвоста, загребая при этом задними ногами, которые были намного больше передних. Когда ранние текодонты вышли из воды на сушу, их сильные задние ноги быстро приспособились к ходьбе по твердой земле.
Вскоре текодонты стали великолепными ходоками и бегунами. Большую часть времени они передвигались по суше на четырех ногах. Однако у них была способность превращаться в истинных спринтеров. Для этого текодонты принимали своего рода "стартовую позу" отклонялись назад, опираясь на свои сверхразвитые задние конечности, и устремлялись вперед на двух ногах, балансируя на бегу длинным хвостом. Ярким представителей этой группы был огромный хищник – пресстозух, он достигал в длину 2-7 м и весил до 700 кг. За последующие 20 млн. лет текодонты развились в первых на Земле динозавров.
Престозух
Фитозавр
Другим отрядом пресмыкающихся, относящийся к надотряду текодонтов подкласса архозавров являлись фитозавры (Phytosauria) - они вели полуводный образ жизни, почти как современные крокодилы. Здесь выделяется семейство pseudopalatinae, включающее в себя крупнейших «почти наземных» животных триаса до 12 м. Третий большой отряд круротарсов – аэтозавры. Эти животные были растительноядными, у них были костные пластины на коже, а некоторые обзавелись и рогами. Типичная длина аэтозавра была 3-5 м.
Динозавры.
Второй большой группе триасовых архозавров являлись орнитодиры. Львиную долю среди них составляют динозавры, которые в триасе только-только появились. Подавляющее большинство триасовых динозавров – ящеротазовые. Самая большая их группа – зауроподоморфы – длинношеие и длиннохвостые растительноядные динозавры с маленькими головами, относятся к отряду прозауроподов, это одна из двух параллельных попыток эволюции сделать огромную живую газонокосилку. В отличие от истинных зауроподов, прозауроподы (кроме самых крупных) сохраняли способность вставать на задние ноги, если потребуется, например, объесть листву с верхушки особо высокого дерева. Крупнейшим семейством триасовых прозауроподов были платеозавриды. Внутри этой группы животных были:
1) мусзавр: Это был один из самых маленьких травоядных животных, которые существовали на планете. У них была длинная шея и хвост, и они были четвероногими. По данным окаменелостей, может достигать до 3 метров в длину и веса около 75 кг.
2) Sellosaurus: также известный как платеозавр. Они были довольно большими, их длина достигала 11 метров, а средний вес - 5 тонн. У них было приблизительно десять шейных позвонков и две пары конечностей, задние ноги были гораздо более развиты и сильнее, чем передние. Что касается их рациона, до недавнего времени считалось, что они явно травоядные, но согласно последним данным у них была всеядная диета.
Платеозавр
Следующим большим семейством триасовых прозауроподов являлись риожазавриды достигавшие примерно таких же габаритов как и платеозавриды.
Отряд истинных зауроподов в триасе только-только формируется (один из самых первых известных его представителей живший 230 млн. лет назад - Эораптор (Eoraptor)), известно лишь четыре рода. Крупнейшие триасовые зауроподы достигали 12 м в длину и 2 тонн веса.
Были в триасе и другие зауроподоморфы – примитивные и переходные формы, в основном мелкие (10-70 кг) и совсем не похожие на зауроподов.
Другая большая группа триасовых динозавров – тероподы появившиеся вместе с зауроподами примерно 230 млн. лет назад старейший из которых является Эодромэеус (Eodromaeus). Это единственный отряд хищных динозавров, все другие динозавры были растительноядными. Самое большое надсемейство хищных триасовых динозавров – целофизоиды, это были очень быстрые и подвижные страусообразные твари длиной от 1,2 до 5,15 м вместе с хвостом и весом до 130 кг. У них были длинные извилистые шеи, острые зубы, когтистые руки и длинный костлявый хвост. Окаменелости целофизов, найденные в большом количестве, указывают на то, что животные охотились группами. Некоторые из крупных обнаруженных окаменелостей имели в брюшной полости остатки меньших членов своего вида. Ученые не могут объяснить, указывает ли это на беременность или, возможно, каннибализм.
Более совершенное строение скелета это основная причина успеха развития динозавров, но всего лишь одна из них. Кроме этого водоотталкивающая кожа динозавров была чешуйчатая и хорошо защищала их от хищников. Яйца динозавров имели прочную скорлупу и поэтому процент выживания детенышей, был очень высок.
Водные рептилии.
Многие животные триасового периода вновь вернулись в морские глубины и стали грозными морскими хищниками. Немногочисленные стегоцефалы, которые выжили после великого пермского вымирания, в результате потепления вынуждены были вновь вернуться в воду, превратившись в грозных хищников. Но главными водными властелинами триасового периода были все же рептилии. Самой большой их группой были завроптеригии – существа, перешедшие к водному образу жизни.
Нотозавр
Рассмотрение завроптеригий начнем с нотозавров имевшие размеры от 18 см до 4 м в длину. Они имели длинное обтекаемое тело, тонкую шею и хвост и охотились исключительно на рыбу. У него была вытянутая морда, в которой было большое количество острых зубов, подходящих для захвата добычи. Несмотря на жизнь в воде, нотозавр выходил на берег загорать.
Другая интересная группа завроптеригий – плакодонты, пытавшиеся занять экологическую нишу морских черепах. К современным черепахам они не имеют никакого отношения, это просто внешнее сходство. В длину плакодонты достигали от 90 см до 2 м. Не у всех плакодонтов панцирь был хорошо развит, у некоторых имелись лишь отдельные костные пластины, почти незаметные. Крупные плакодонты были похожи на тритонов и ползали по морскому дну, выискивая раковины, а затем раздавливали их мощными плоскими зубами.
В водах триасового периода процветали различные виды ихтиозавров. Ископаемые останки ранних ихтиозавров похожи на ящериц, их позвонки указывают, что они, вероятно, плавали, перемещая свои тела в сторону, как современные угри. В дальнейшем шейные позвонки ихтиозавра постепенно срастаются в одну кость. Ещё позже ихтиозавры превратились в чисто морские формы с дельфинообразными телами и длиннозубыми мордами. И уже к середине триаса ихтиозавры доминировали в океанах. Эти четвероногие, подобно современным дельфинам и китам, окончательно перешли к полностью водному образу жизни, приобрели гидродинамически совершенную форму и освоили живорождение. Пятипалые конечности ихтиозавров превращаются в хорошо приспособленные для плавания ласты. Внешне они стали очень похожи на дельфинов, единственное заметное отличие – задние ноги. У ихтиозавров они превратились в брюшные плавники, у дельфинов - атрофировались. У ихтиозавров осталось легочное дыхание. В отличие от китообразных, ихтиозавры не пользовались эхолокацией, а полагались в основном на зрение. Из-за этого их глаза были огромны, у некоторых видов – до 20 см в диаметре. Самое большое семейство триасовых ихтиозавров – шастазавры. Самый крупный из них - шонизавр. Он был самым большим ихтиозавром всех времен и достигал в длину 23 м, а весил до 40 тонн, это был абсолютный рекорд размеров для животных, предыдущий рекорд (акула камподус каменноугольного периода) превышен почти вдвое. Другие шастазавры, впрочем, были гораздо мельче.
Еще одна группа рептилий, появившиеся в триасе и делившая морскую среду обитания с другими видами, называется плезиозавры. Они были сравнительно невелики, до 2 м в длину. В следующем, юрском периоде, плезиозавры развились до гигантских размеров. Они обитали в море, но иногда выходили на берег. Питались рыбой.
Летающие рептилии.
В конце триасового периода в эволюции жизни на Земле произошли два наиважнейших события. Одно из них случилось на суше и ознаменовалось появлением первых млекопитающих. Второе произошло в воздухе, и было связано с пришествием птерозавров ("крылатых рептилий").
Некоторые позвоночные животные уже пытались подняться в воздух. Одним из "первопроходцев" была небольшая ящерица, жившая в пермский период и названная учеными вешельтизавр. Однако у нее не было настоящих крыльев. Она планировала с дерева на дерево на своего рода перепончатом крыле, натянутом между невероятно длинными ребрами.
Но на смену им быстро пришли птерозавры, вместо ребер переделавшие под полет структуру конечностей, между которыми образовалась специальная воздухоплавательная пленка, при помощи которой они могли, планируя в воздушных потоках, подолгу оставаться в воздухе, с высоты высматривая себе добычу. В отличие от современных летучих мышей, у птерозавров летательная перепонка держится не между растопыренными пальцами руки, а между мизинцем руки и ногой, это обеспечивает лучшие летные качества, особенно при планирующем полете, но усложняет перемещения по земле со сложенными крыльями. Размах крыльев триасовых птерозавров составлял 45-75 см. Первые птерозавры жили на берегах рек и питались насекомыми. Они были яйцекладущими и имели длинный клюв.
В триасе появился особый представитель триасовых архозавров - протоавис, предок всех птиц. Впрочем, не все палеонтологи согласны с этим утверждением.
Птерозавр
Протоавис
Млекопитающие.
Теперь перейдем к «млекопитающей» ветви эволюции рептилиоморфов – синапсидам. Пермско-триасовое вымирание сильно проредило их ряды. Самая распространенная группа синапсид – цинодонты — быстроногие хищные рептилии, нападавшие на стада медлительных растительноядных животных. Эти звери внешне практически неотличимы от примитивных млекопитающих. Кратко перечислим их основные семейства в порядке убывания родового разнообразия. Маленькие (10-15 см) насекомоядные probainognathidae, traversodontidae – умеренно крупные (до 1,8 м) растительноядные, Cynognathia – небольшие собакоподобные хищники (50см - 1 м), Galesauridae – самые примитивные цинодонты, возможно являвшиеся предками всех остальных, Tritylodontidae – растительноядные наподобие современных крыс достигавшие 50 см в длину.
Цинодонт
В самом начале триаса кратковременно распространились более примитивные дицинодонты – одна из ветвей аномодонтов, в перми это были такие маленькие свинки с двумя длинными зубами, торчащими из верхней челюсти. В триасе дицинодонты сильно выросли, в длину они составляли 2-4 м, а по массе – от 400 кг до 2 т. Однако когда появились крупные круротарсы, звезда дицинодонтов закатилась, они продолжали существовать до конца мезозоя, но уже как единичные виды.
В конце триаса цинодонты дали начало настоящим млекопитающим, с грудиной, диафрагмой и молочными железами. Эти наши далекие предки были мелкими (10-12 см) зверьками, похожими на современных мышей и землероек. Это древнейшее млекопитающее называется дроматериумом, что в переводе значит “бегающий зверь”. Дроматериум подобно современным австралийской ехидне и утконосу, не рождали детенышей, а откладывали яйца, из которых вылупливались недоразвитые детеныши. В отличие от пресмыкающихся, совершенно не заботившихся о своих потомках, дроматериумы выкармливали детенышей молоком.
В процессе эволюции из дроматериума развилась новая ветвь животных, которых назовут млекопитающими. Как правило, они были травоядными или насекомоядными, и поэтому не находились в прямой конкуренции с архозаврами или динозаврами. Многие из них, вероятно, вели частично древесный и ночной образ жизни. Большинство из них явно имели шерсть. Согласно исследованиям, эти животные относились к теплокровным. На протяжении последующих 160 миллионов лет, млекопитающим было суждено находиться в тени динозавров.
Растительность.
Растительный мир суши наследовал черты позднепермской эпохи. Подводная растительность этого времени не перетерпела изменений – водоросли, характерные для пермской эпохи, продолжали процветать. Все также прогрессировали сине-зеленые, бурые и прочие виды водорослей, характерные еще для палеозоя. Большое значение на данный период оказывали рифообразующие водоросли.
Не претерпели значительных изменений по сравнению с предшествующей ему пермью и наземные растения. Растительный мир суши 1-й половины триасового периода во многом близок к верхнепермскому; в тропиках преобладают, ныне вымершие, птеридоспермы и древние хвойные, в умеренной зоне господствуют разнообразные палеофитные папоротники. На всех континентах широко распространены своеобразные плауновидные, специфические для триасового периода. Хотя к середине триасового периода вследствие сглаживания критических различий между погодными условиями различных климатических поясов, заметного разделения растительных видов по всей территории Пангеи не прослеживается.
Основные мезофитные группы растений (диптериевые папоротники, саговники, беннеттиты, гинкговые, мезофитные хвойные) в значительном количестве появляются во 2-й половине триасового периода, но вплоть до конца триасового периода ещё велико значение древних групп. Некоторые саговниковые дожили и до наших времен. Встречаются они в зоне Малайского архипелага и называются саговыми пальмами. По своей сути саговники являются промежуточной ветвью развития между папоротниковыми и пальмами. Они, так же как и пальмы, имели мощный ствол, разветвленную пальмовую перистолистную крону наверху, но размножались все еще не семенами, а микро или макроспорами. А поскольку их споры были наихудшим образом защищены от холодов, выжить данные растения могли только в тех континентальных зонах, которые никогда не смещались к зонам холодов и постоянно были поблизости от воды.
В позднем триасе исчезли около половины всех наземных растений. Исчезли древовидные плауновые и каламитовые, кордаитовые, прапапоротники и большая часть древних хвойных.
Триасовая катастрофа.
Конец триасового периода ознаменовало новое массовое вымирание видов, сравнимое с аналогичным событием в конце перми. Его причины остаются загадкой. Вот гипотезы о причинах, которые привели к исчезновению видов:
- Падение метеорита. Против данной теории свидетельствует то, что не обнаружен кратер от астероида. Маникуаганское водохранилище, предположительно считающиеся местом падения метеорита, возникло за 12 млн. лет до массовой гибели животных;
- Изменение климата. Возможной причиной вымирания называют наступление ледникового периода. Высказывалась гипотеза о том, что произошло изменение уровня воды в океане. Но эта версия не объясняет гибель подводных обитателей;
- Повышенная вулканическая активность. Вулканические выбросы диоксида серы и углекислого газа влияют на климат и способны вызвать глобальное потепление или похолодание. В почве были обнаружены доказательства повышения уровня углекислого газа в период триасово-юрского вымирания;
- Понижение уровня кислорода в океане.
Биологическая катастрофа унесла жизни 22% семейств морской фауны и 52% морской флоры [море "задохнулось"]. Вымерли:
- оставшиеся роды палеохиноидей (семейство Perischoechinidae),
- род ортоцеровых (Orthoceras),
- большинство триасовых семейств и родов аммонитов и цератитов (Medicottidae, Ceratitidae, Ptychitidae, Pinacoceratidae, Tropitidae, Cyclolobidae, Arcestidae, Cladiscitidae),
- многие группы пресмыкающихся: Parasuchia, Placodontia, Catylosauria, Pelycosauria, Theriodontia (зверозубые), Anomodontia, Rhynochosauria.
Синхронно с великим морским вымиранием, происходит и падение разнообразия насекомых. Вымерли многие наземные животные. Гибель происходит быстро – за 10.000 лет. В целом, триасовое вымирание стёрло с лица планеты почти половину видов и открыло дорогу взлёту динозавров.