В предыдущей статье мы рассмотрели, при помощи учебника, из каких элементов состоит ДНК и выяснили, что в состав молекулы входит всего 6 типов небольших молекул: дезоксирибоза, азотистые основания (аденин, тимин, гуанин и цитозин) и фосфат. В параграфе “Макромолекулярная структура ДНК” рассказывается, как все эти молекулы и составляют главную молекулу ДНК.
“В 1953 г. Дж. Уотсон и Ф. Крик предложили модель структуры ДНК. При построении модели ученые основывались на четырех группах данных:
1. ДНК представляет собой полимер, состоящий из нуклеотидов, соединенных 3'-5'-фосфодиэфирными связями.
2. Состав нуклеотидов ДНК подчиняется правилам Чаргаффа: в любой ДНК содержание пуриновых нуклеотидов (А+G) всегда равно содержанию пиримидиновых нуклеотидов (Т+С); число остатков А всегда равно числу остатков Т, число остатков G — числу остатков С.
3. Рентгенограммы волокон ДНК, впервые полученные М. Уилкинсом и Р. Франклин, указывают на то, что молекула обладает спиральной структурой и содержит более одной поли нуклеотидной цепи.
4. Кислотно-щелочное титрование ДНК показывает, что ее структура стабилизируется водородными связями. Титрование и нагревание нативной ДНК вызывает заметные изменения ее физических свойств, в частности вязкости, переводя ее в «денатурированную» форму, причем ковалентные связи при этом не разрушаются.
На основании этих данных Д. Уотсон и Ф. Крик предложили трехмерную модель ДНК, которая объясняла результаты рентгеноструктурного анализа и характерную для ДНК парность оснований (рис. 42).” (Стр. 83-84).
Ученые получили такую схематическую модель молекулы.
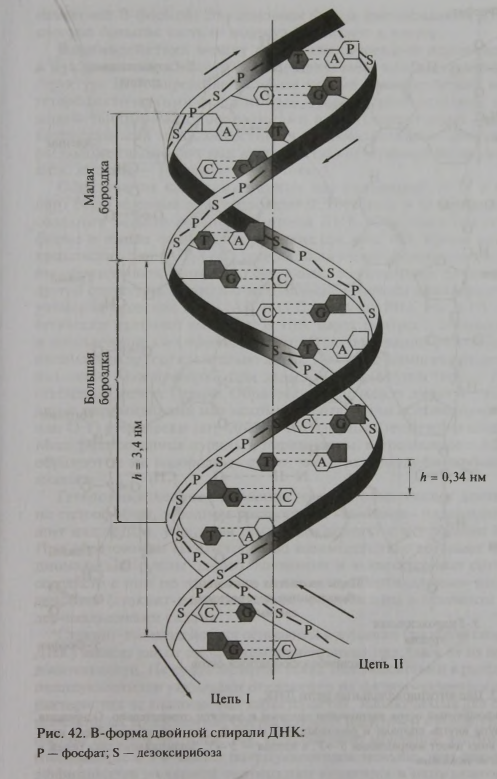
Я видел в каком-то ролике, что они построили такую вертикальную модель из разноцветных шаров выше самого Уотсона. Для нас важным в этой модели является то, что если, двигаясь по одной из цепей в одном направлении, мы обнаружим тимин (T), то парой ему будет обязательно аденин (A). Не гуанин (G), не цитозин (C), а именно аденин (A). Если на этой же ветви дальше встретим цитозин, то парой ему будет гуанин и не что иное. Для аденина парой будет тимин, а для гуанина парой будет цитозин. Такие пары называются комплементарными.
Таким образом, если мы разорвем цепи и определим какие основания и в каком порядке есть на одной из них, то без труда определим какие основания и в каком порядке следуют основания на другой цепи. Это и дает природе возможность по одной цепи достроить верную вторую цепь. То есть, разделив одну ДНК на две цепи, а затем достроить к каждой из цепей вторую цепь, можно получить две совершенно одинаковые ДНК. Этот процесс называется репликацией. Так из одной молекулы можно получить сколь угодно много реплик совершенно одинаковых ДНК.
Все это верно, но на этом закончился гений Д. Уотсона и Ф. Крика, и за этим последовала не объяснимая бессмысленность. В чем она заключается мы и рассмотрим в последующих статьях.