Квантовая биология не отрицает наличия всех фаз клеточного деления, тем более что эти фазы наблюдаемы при помощи различных инструментов, но интерпретацию некоторых этих событий предлагает иную.
Многие эксперименты показывают, что центриоли есть в половых клетках. Казалось бы, что центриоли передаются по наследству, но тот же опыт показывает, что это не так. В центриолях не находят достаточного количества генетического материала. Очень изредка обнаруживаются элементы ДНК, и намного чаще обнаруживается РНК, но этого недостаточно для процесса репликации центриоли. Да и вновь синтезируемая центриоль никак не связана с материнской центриолью.
Вот что пишет Узбеков:
У насекомых в процессе раннего развития, подробно исследованного на примере дрозофилы, в течение первых 2.5 ч синцитиальные деления проходят примерно каждые 10 мин, в результате образуется несколько тысяч центриолей. Этот процесс не зависит от транскрипции, т. е. все эти центриоли образуются из материалов, накопленных в ооците. Таким образом, передача триплетной структуры через центриоли базального тельца сперматозоида не имеет решающего значения, а отцовские центриоли имеют не генетическое, а лишь организующее значение. (стр. 106).

И так, мы имеем клетку с двумя центриолями, изображенными на Рис. 1.
Опыт показал, что микротрубочки, которые входят в состав центриолей, почти такие же, как и микротрубочки, тянущиеся от центриолей до стенок клетки и даже иногда прорастающие через стенку клетки в виде жгутиков и ресничек во внешний мир. Рис.2.
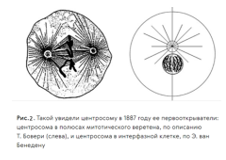
На втором рисунке изображена центросома, в которой и содержатся центриоли, а лучики – это микротрубочки. Они и удерживают центросому на месте, в большинстве случаев в центре клетки.
Как же начинается процесс деления клетки? Узбеков по этому поводу пишет, что в это время ”…запускается сложная цепь биохимических реакций…”. А именно. ЦиклинD – CDK 4,6 фосфолирует белок pRB, который освобождает факторы активации транскрипции EF2, а эти факторы активизируют синтез циклинов E и A, что и приводит к началу дупликации центриолей.
В статье о репликации я писал, что процесс репликации запускается внешними тепловыми фотонами непосредственно или через некоторую цепочку, полагаю не более двух звеньев. Узбеков предлагает 5 ступеней: циклин-белок, белок-факторы, факторы-циклины, циклины-дупликация. 4 ступени, но еще надо подать какой-то сигнал циклинуD, чтобы он заработал. Всего 5 ступеней.
Похоже, что и клеточный цикл провоцируют тепловые фотоны. Может быть те же, которые запускают репликацию, а может быть и разные. Это можно определить, если получится запустить деление клетки без репликации или запустить репликацию без деления клетки.
Узбеков согласен, что и репликация ДНК, и синтез центриолей осуществляется одним и тем же: ”…оказалось, что оба процесса – репликация ДНК и удвоение центриолей – регулируются одними и теми же цитоплазматическими механизмами”. (стр. 98).
И это, не смотря, на то, что: ” В настоящее время показано, что репликация центриолей может начаться в G1-фазе, т. е. До начала репликации ДНК” (Узбеков, 2007). (стр. 99).
Сигнал на репликацию подан. ДНК начала делится, связи между основаниями начали плавиться, как льдинки при 0 градусов. Точно также начали разрываться связи между молекулами тубулина в микротрубочках. В клетке появился свободный тубулин. Но МТ в центриолях имеют дополнительные связи между собой и окружены перицентриолярным материалом, и они не распадаются.
Свободный тубулин, также, как и молекулы воды, воска, атомов металлов и другое, в точке бифуркации самостоятельно соединяется и разъединяется из-за тепловых флуктуаций. И если в дополнение к этим спонтанным фотонам добавить еще какой-нибудь поток фотонов, той или иной энергии, то тубулин будет либо активней кристаллизоваться, либо вообще не будет соединяться.
Уцелевшая центриоль и является таким катализатором, который под воздействием одной из митохондрий исходной клетки, генерирует свет, приводящий к началу синтеза новой центриоли. Но этот свет (фотоны определенной энергии) каждый по отдельности не способен катализировать реакцию между молекулами тубулина, а вот совместное их воздействие на узел связи между молекулами позволит молекулам образовать устойчивую связь.
И вот оказалось так, что эти совместные сгустки энергии (фотоны), генерируемые центриолью, располагаются в плоскости параллельной к данной центриоли. И как только в это поле попадает тубулин он сразу же соединяется с такой же другой молекулой. А это голографическое поле получилось в точь, как поперечный разрез центриоли. Образовалось, так называемое, “тележное колесо”. Дальше на это колесо начало нарастать и молекулами тубулина, и всевозможными молекулами связи МТ, и всем остальным по мере поступления материала, пока не распадутся все исходные МТ.
Поскольку центриоль обладает осевой симметрией, то колес может образовываться несколько. Это отчасти случайный процесс. Вот что пишет поэтому поводу Узбеков:
” При исследовании ресничного эпителия удалось обнаружить структуры, в связи с которыми в клетках формируются центриолярные цилиндры – дейтеросомы, от каждой из которых может одновременно расти от 1 до 14 процентролей (Sorokin, 1968). Достигая половины длины, процентроли отделяются от дейтеросом и продолжают расти до длины зрелой центриоли. Сформировавшиеся центриолярные цилиндры мигрируют к клеточной поверхности…” (стр. 98).
Соответственно это способствует ” …появлению многополюсных митозов (Мэзия, 1963). (стр. 99).<.p>
Дальше, как описывает Узбеков, происходит следующее:
В конце интерфазы или в профазе митоза две пары центриолей начинают расходится и формируют два равнозначных центра полимеризации МТ – профазные “звезды”, при этом интерфазные МТ полностью разрушаются. (стр. 97).
Действительно с этим легко согласится. Центриоли освободились от внешних МТ и если, например, материнские центриоли обращены друг к другу своими минус-концами, а дочерние центриоли расположены своими однополюсными концами в одну и ту же сторону, то такие пары будут отталкиваться друг от друга и разойдутся к полюсам.
Теперь каждый центр будет достраивать свои центриоли и строить на них микротрубочки. Весь наличный в клетке материал будет расходоваться на строительство этих элементов. Если материала много, то микротрубочки могут дотягиваться до реплицирующейся ДНК, и, как полагают ученые, закрепятся на центромерах хромосом. И даже обязаны закрепиться на хромосомах, чтобы потом эти микротрубочки растащили, к каждому центру именно свои хромосомы. Получилось веретено.
Все хорошо пока не доходишь до таких рассуждений “Белки-моторы, ассоциированные с МТ, участвуют в процессе формирования митотического веретена и осуществляет разнообразный транспорт вдоль МТ интерфазной сети. При этом МТ выступают в качестве своеобразных рельсов, по которым органеллы и белковые комплексы перемещаются в обоих направлениях”. (стр. 102).
Дальше говорится, что от центра клетки к периферии бегают белки кинезины, а в противоположном направлении движутся белки денеины. И что? на каждый груз требуется один кинезин или денеин? Или они бегают обратно порожняком? И какие такие грузы перемещаются по этим рельсам?
Увы, никаких таких транспортных устройств в клетке нет. Да они и не нужны с точки зрения квантовой биологии. А с точки зрения молекулярной биологии они необходимы, ибо как клеточный центр притащит к себе хромосомы. Последние достижения в познании процесса разделения хромосом и описаны в статье “Полчаса митоза”.
Наш ученый Фазли Атуаллаханов сделал в Америке (нам он не нужен) лазерный пинцет, которым можно работать с хромосомами, микротрубочками и другими мелкими объектами. И вот, что он сделал с американскими учеными:
“Когда ученые из Стэнфорда Д. Друбин и Дж. Барнс выделили белки, из которых в их опытах собиралось колечко на микротрубочке, исследователи митоза страшно обрадовались. Сложилась вполне логичная гипотеза: микротрубочка не просто утыкается в хромосому — на ней образуется кольцо, к кольцу цепляется «висюлька». После разрезания (хромосом – авт.) концы трубочки выгибаются и давят на кольцо. Оно под давлением начинает съезжать по трубочке и тащить за собой хромосому. Но это были лишь теоретические построения, проверить которые не позволяли ни микроскопы, ни биохимические методы, ни рентгеноструктурный анализ. Пока не появился лазерный пинцет”.
Примерно как на рисунке 3.
Микротрубочка разрушается в виде воронки, и этим тащит колечко за собой, а колечко через висюльку тянет хромосому. И это так у них получилось на опыте. Единство теории и практики на лицо. Но эту идиллию немножко портит то событие, что иногда трубочка тянет хромосому, не приходя в соприкосновение с ней. Перед учеными встала задача найти механизм, тянущий хромосому без колечка. Пока что-то не слышно, чтобы эта проблема была решена, да и вряд ли ее можно решить на молекулярном уровне.
На квантовом уровне эта задача решается просто. После того, как клеточные центры разошлись, они начали строить микротрубочки внешние по отношению к центриолям и внутренние. Но что значит строить? Некоторые элементы (электроны в молекулах) излучают свет (фотоны соответствующей энергии), которые являются резонансными для некоторых электронов в других молекулах. В частности, в молекулах тубулина и ДНК. Как это происходит описано в статьях "Спонтанное излучение" и "Вынужденное излучение" . Что значит резонансные фотоны – это те фотоны, которые способны поглощаться электронами и под воздействием возникающей в результате поглощения силы, двигаться навстречу движения фотона. Это обычная гравитация.
Этого не следует бояться. Особенно если вы верите в дуализм и суперпозицию, которые ничего не объясняют, кроме опыта Юнга, и то неверно, то легче поверить в гравитацию, которая объясняет очень многое.
И так. Клеточный центр стягивает к себе все, что поглощает его излучение. Первым делом это тубулин. Где бы не находилась его молекула, он будет привлечен к той или иной трубочке. Вступит с ней в реакцию. Если природой предусмотрено, что для организации морфологии МТ потребуется еще какая-то связь, то соответствующий элемент этой связи либо сам притянет к себе контрагента, либо контрагент своим светом подтянет к себе тубулин. Возможно этот тубулин организует связь с соседней трубочкой или еще что-нибудь подобное.
Как только молекула тубулина (или другого вещества) завершит свои внутренние дела, она снова засверкает и будет требовать для закрытия этого света следующую молекулу. Этот свет будет стремится даже подтянуть хромосому, но в этом случае у него ничего не получится – для этого случая хромосома слишком тяжела. И снова придется довольствоваться молекулой тубулина.
Такие события будут происходить с каждой МТ до тех, пор пока в клетке будет свободный тубулин. Hо вот запасы тубулина начинают заканчиваться. Не удовлетворенных (светящихся) связей оказывается все больше и больше. Но единственный объект, в котором есть резонансные электроны для света МТ – это хромосомы. Они все сосредотачиваются на них. И, наконец, наступает период, когда совместных гравитационных усилий микротрубочек становится достаточным, чтобы разорвать хромосому. Ну а дальше разрывается вся цепочка хромосом, как начавшийся рваться лист бумаги или лоскут материи в ваших руках после первого надрыва.
По пути движения молекула ДНК удовлетворяет связи некоторых, которые первые встречаются, МТ, но остальные трубочки тянут молекулу к центру, разрывая образовавшиеся связи, которые снова открывают свои резонансные свойства. Движение ДНК продолжается, пока определенное количество МТ с возбужденными связями не окажется позади ДНК, и они не начнут тормозить движение молекулы. При равновесии этих сил молекула и остановится. Остановка по воле судьбы оказывается вблизи клеточного центра.
Не закрытые связи микротрубочек все просят своего удовлетворения, провоцируя (экспрессируя ген тубулина) этим ДНК на синтез тубулина. Они получают свои порции тубулина и продолжают расти. Трубочки от центров растут навстречу друг другу. Потенциалы встречных концов МТ одинаковые и они добавляют свои расталкивающие силы к расталкивающим силам центриолей. Этих сил достаточно, чтобы разорвать, по сути состоящую из липидов, клетку.
Прорванные бреши быстро затянутся благодаря водородным связям липидов и благодаря еще пока отсутствующему избытку давлению в клетке (она дырява).
Наконец МТ уткнутся в места, где закроют возбужденные связи. ДНК начнет генерировать свои внутренние белки и готовиться к следующему делению. Жизненный виток совершился.
На сколько достоверна предложенная квантовая модель митоза я не знаю. Но я уверен, что истинную модель митоза, как всего другого, можно построить только на квантовом уровне с привлечением квантовой физики.