СТАТЬЯ
Открытый Доступ
Геномное происхождение Заны Абхазской
Ашот Маргарян, Миккель-Хольгер С. Синдинг, Christian Carøe, Владимир Ямщиков, Игорь Бурцев, М. Томас П. Гилберт
Впервые опубликовано: 14 июня 2021 года.
https://doi.org/10.1002/ggn2.10051
Информация о финансировании: Этот проект был профинансирован Внутренним грантом Копенгагенского университета MTPG.
Абстрактный
Загадочные явления пробудили воображение людей по всему миру к созданию фольклорных существ. Одним из ярких примеров является Зана из Абхазии (Южный Кавказ), хорошо документированная женщина 19 века, которая была захвачена, живя в дикой природе в лесу. Внешность Заны была достаточно необычной, что местные жители называли ее Альмастой—аналогом снежного человека на Кавказе. Хотя точное место захоронения Заны было неизвестно, могила ее сына Хвита была идентифицирована в 1971 году. Геномы Хвита и предполагаемого скелета Заны были секвенированы на среднюю глубину около 30 см. 3× использование древних методов ДНК. Идентичные мтДНК и отношения между родителями и потомками между ними указывали на то, что неизвестная женщина действительно была Заной. Популяционный геномный анализ показал, что непосредственные генетические предки Заны, вероятно, могут быть прослежены до современных восточноафриканских популяций. Мы предполагаем, что у Заны могло быть генетическое расстройство, такое как врожденный генерализованный гипертрихоз, который мог частично объяснить ее странное поведение, отсутствие речи и длинные волосы на теле. Наши находки проясняют печальную историю Заны и дают ясный пример того, как предрассудки того времени привели к появлению представлений о загадочных гоминидах, которые до сих пор сохраняются и передаются некоторыми сегодня.
1 ВВЕДЕНИЕ
В местном фольклоре южнокавказского региона Абхазии упоминается “дикая женщина” по имени Зана, жившая в 19 веке, которую некоторые местные жители называли самкой Абнауаю или Алмасты: так называют существо, похожее на печально известного йети Гималаев и снежного человека Северной Америки , предположительно обитающего на Кавказе и в Средней Азии. Первоначально захваченная в плен во время жизни на открытом воздухе в лесу, Зана позже была порабощена чередой местных богатых людей и, наконец, куплена абхазским дворянином Эджи Генабой, который отвез ее в свое поместье в Тхине, где она жила до своей смерти около 1890года.Вдохновленные предположением, что она могла быть самкой йети, советские ученые посетили регион в 1962 году, чтобы собрать описания и отчеты от старейшин, живущих в деревне Тхина, которые все еще помнили ее. Местные жители описывали ее как “наполовину человека, наполовину животное”, 2 м ростом, темнокожую, покрытую густой шерстью, которая могла одной рукой поднять 50-килограммовый мешок муки и обогнать лошадь в беге.1, 3 По свидетельствам очевидцев, ей также не хватало речи, что наряду с ее якобы странным поведением и внешностью, вероятно, привело к ее репутации Альмасты. Зана также родила двух сыновей и двух дочерей от местных мужчин. После ее смерти она была похоронена на семейном кладбище Генабас, и хотя точное место захоронения Заны было неизвестно, могила ее младшего сына Хвита была идентифицирована в 1971 году. После нескольких попыток найти место захоронения Заны на семейном кладбище Генабы были обнаружены останки неизвестной женщины, что привело к предположению, что они , возможно, принадлежалисамой Зане.Чтобы преодолеть ограничения предыдущего анализа ДНК и возможные неясности краниометрических исследований, мы секвенировали геномы как неизвестной самки, так и Хвита до 3,1 - и 3,3-кратного охвата соответственно. Мы провели геномный анализ, чтобы исследовать их генетическую родословную и родственные отношения, что позволило нам пролить свет на историю Заны, основанную на объективных геномных данных.
2 МАТЕРИАЛ И МЕТОДЫ
См. Дополнительные методы для более глубокого описания материалов и методов, используемых в этом исследовании.
2.1 Генерация данных и биоинформатический анализ
Все лабораторные исследования древней ДНК (аДНА) проводились в специальных чистых лабораторных помещениях Института ГЛОБУСА Копенгагенского университета в соответствии со стандартами аДНА, описанными в других источниках.6Мы использовали зубы и окаменелые кости для извлечения древней ДНК из обоих индивидуумов и создания двухцепочечных библиотек BGISeq в соответствии с ЛУЧШИМ протоколом, используя адаптеры, совместимые с секвенированием BGI в соответствии с Mak et al. 2017.7 Усиленные библиотеки были секвенированы на двух полосах платформы BGISEQ-500.Мы использовали рабочий процесс BAM, реализованный в конвейере PALEOMIX pipeline8, чтобы обрезать и сопоставить результаты секвенирования с эталонной сборкой генома человека GRCh37 и пересмотренной эталонной последовательностью Cambridge reference sequence (rCRS, NCBI accession number NC_012920.1).Мы использовали mapDamage v2.0, чтобы получить распределение длины считывания и приблизительные байесовские оценки параметров повреждения.9 Для оценки уровней загрязнения в древних образцах мы использовали contamMix10 и метод заражения на основе Х-хромосомы, реализованный в ANGSD.11 Пол индивидуумов был определен в соответствии с оценками Ry, описанными в другом месте.12
2.2 Анализ однопартийных маркеров и родства
Мы использовали haplogrep213 для назначения гаплогрупп мтДНК. Для определения гаплогруппы Y хромосомы Хвита мы использовали рабочий процесс pathPhynder (https://github.com/ruidlpm/pathPhynder).Филогенетический сетевой анализ полных последовательностей мтДНК, в том числе последовательностей “Zana”, Khwit и других гаплогрупп L214 (n = 93), проводился с помощью POPART15 по алгоритму “Медианная соединительная сеть”. Мы использовали BEAST v2.6.116 для оценки времени дивергенции линии мтДНК “Zana” с использованием метода байесовской линии горизонта (BSP). Высота дерева была откалибрована на основе предыдущей работы Silva et al.14Коэффициенты родства были рассчитаны путем первого создания файла правдоподобия частоты аллелей сайтов (saf) в ANGSD (http://www.popgen.dk/software/index.php/IBSrelate) , за которым последовала оценка матрицы совместного использования IBS на основе двумерного частотного спектра сайта (2d-SFS) из real-SFS, реализованного в ANGSD.11
2.3 Популяционная генетика
Чтобы оценить генетическую связь между “Заной”, "Хвитом" и другими популяциями, мы объединили данные секвенирования дробовика от исторических особей с панелью массивов SNP Affymetrix Human Origins мировых популяций.17, 18 Мы также включили для сравнения данные трех архаичных людей (два неандертальца19 и денисовец20) и генотипов шимпанзе, а также геномы двух кавказских охотников-собирателей (CHG) (образец SATP возрастом около 13300 лет и образец KK1 возрастом около 9700 лет), первоначально раскопанных на Южном Кавказе21.Мы провели неконтролируемый кластерный анализ на основе максимального правдоподобия с ПРИМЕСЬЮ22, обрезав набор данных для неравновесия связей с помощью plink v1.9.23 Программа pong24 использовалась для определения и визуализации наилучшего прогона для каждого K и аналогичных компонентов между различными K.Анализ главных компонентов (PCA) проводился с использованием plink v1.9 с проецированием древних геномов на современную вариацию. Первые 30 собственных векторов PCA были использованы в качестве входных данных для анализа равномерной аппроксимации многообразия и проекции (UMAP) с использованием R-пакета “umap”.Оценки D-статистики рассчитывались с использованием пакета ADMIXTOOLS25 и R “admixr” 26. Максимально вероятное филогенетическое дерево “Зана” и африканских популяций было построено спомощью Treemix27.
3 РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В общей сложности 1 219 599 801 последовательность BGISeq была сгенерирована из зубов и окаменелых костей обоих индивидуумов (см. Картографическую статистику в таблице S1). Как и следовало ожидать из результатов предыдущих исследований,28 эндогенное содержание было значительно выше в костях петрушки (“Зана”-41,95%, Хвит—33,93%), чем в образцах зубов (“Зана”-1,16%, Хвит—12,7%). Мы использовали полученные данные для получения геномов со средней глубиной секвенирования покрытия 3,1× и 3,3× для “Зана” и Хвита соответственно. Эти последовательности показывают типичные профили повреждений древней ДНК и короткие длины фрагментов ДНК[29]. несмотря на то, что особи умерли относительно недавно, Зана жила примерно до 1890 года, а Хвит-до 1954 года.Оценки загрязнения, основанные на мтДНК (для обоих) и Х-хромосоме (только для Хвита), составляли менее 1%, а хромосомные полы индивидуумов соответствовали их антропологическим описаниям: Хвит-мужчина, а “Зана” - женщина.
3.1 Одноплодные маркеры
Последовательности мтДНК были идентичны для обеих особей, что согласуется с гипотетическими отношениями матери и сына, и могут быть отнесены к гаплогруппе L2b1b, родительская гаплогруппа которой (L2b clade) широко распространена в Западной Африке,14, 30, но также встречается повсей Африке31. Та же гаплогруппа была идентифицирована более ранним, независимым анализом образцов зубов из Хвит и “Зана” в Южном научно-исследовательском институте на основе обогащенных библиотек ДНК человека (неопубликованные данные).Y-хромосомная линия Хвита принадлежит к гаплогруппе R1b1a1b1b, что явно отражает его неафриканское отцовское наследие. Это субгаплогруппа крупной гаплогруппы R1b1a1b, определяемая мутацией M269, которая встречается на высоких частотах в Европе и Западной Азии.32-34Хотя более раннее исследование, в котором анализировалась последовательность мтДНК Хвита, показало, что материнское происхождение "Заны" было африканским,35 авторы предположили, что "Зана" могла принадлежать к древней африканской линии, вероятно, из-за отсутствия подходящего всеобъемлющего набора сравнительных данных в то время. Поэтому мы провели анализ митохондриальной сети, чтобы оценить связь предполагаемой материнской линии образца “Zana” с другими (n = 93) последовательностями гаплогруппы L2 человека14, и обнаружили, что она кластеризуется вместе с другими особями линии L2b, как и ожидалось (рис. 1).
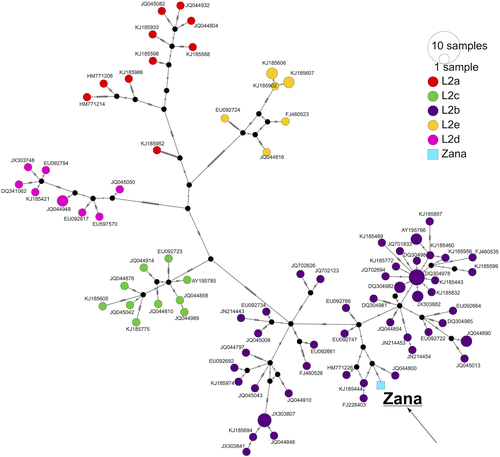
РИСУНОК 1Откройте в программе просмотра рисунковPowerPointМедиана-соединительная сеть. Анализ последовательностей мтДНК “Zana” и “Khwit "наряду с 93 полными последовательностями мтДНК из митохондриального клада человека L2 с использованием алгоритма" Медиана-Присоединение”, реализованного в PopArt. Каждый кружок представляет определенный гаплотип; меньшие черные кружки обозначают медианные векторы. Маленькие черные линии, соединяющие ветви между гаплотипами, обозначают количество шагов мутации, разделяющих гаплотипы. Поскольку последовательности мтДНК “Zana” и Khwit идентичны, в сюжете упоминается только гаплотип мтДНК Zana
Сильва и др. мы подсчитали, что митохондриальная клада L2b, вероятно, возникла около 24 тыс. лет назад,14 таким образом, наше назначение мтДНК может быть использовано для отклонения гипотезы о том, что гипотетический образец “Зана” имел древнее или архаичное происхождение. Чтобы получить приблизительную оценку времени до самого последнего общего предка (TMRCA) материнской линии “Zana” и ее сестринских групп, мы провели анализ ЖИВОТНЫХ только на основе последовательностей L2b (n = 57) (рис. S3). При ограниченном числе последовательностей L2b1b (n = 4) в наборе данных время дивергенции оценивалось примерно в 9800 лет (3515-13 000; 95% наибольших интервалов задней плотности).
3.2 Родство
Далее мы еще проверили, действительно ли два человека были напрямую связаны с использованием ПК-относятся как это реализовано в PCAngsd36 и realSFS.11 родство коэффициент 0.1818 был получен для пары “Зана” и Khwit, таким образом, подпадающие первая степень родства интервал (0.177-0.354), как описано в Manichaikul и соавт.37 Значение R0 (R0 = 0,0015), близкое к 0, было оценено на основе анализа realSFS, предполагая связь родитель-потомство, которая вместе с идентичными мтДНК указывает на то, что неизвестный женский скелет был матерью Хвита, таким образом, может быть положительно идентифицирована как Зана.
3.3 Генетическое сходство
Для дальнейшей оценки генетической связи между Zana, Khwit и различными мировыми популяциями с использованием вариации ядерного генома мы провели PCA, основанный на группе происхождения человека (HO). Чтобы облегчить визуализацию, мы сократили общее количество референтных популяций в панели, чтобы представить основные генетические линии мира. Однако, учитывая его географическую значимость, мы включили относительно больше популяций с Кавказа и, учитывая предыдущие гипотезы о том, что Зана, возможно, имела какое-то архаичное происхождение гоминидов, мы также включили данные генома трех архаичных людей и использовали шимпанзе в качестве аутгруппы. Кроме того, мы включили для сравнения двух мезолитических охотников-собирателей (SATP и KK1) с Южного Кавказа.Результаты ясно показывают, что Зана генетически не близка ни к архаичным людям, ни к шимпанзе, но тесно связана с современными человеческими популяциями (рис. 2А). Как и следовало ожидать из анализа родства (родитель-потомство) и Y-хромосомы (европейская линия R1b1a1b), Khwit занимает промежуточное положение на участке PCA между европейскими или кавказскими и африканскими популяциями.
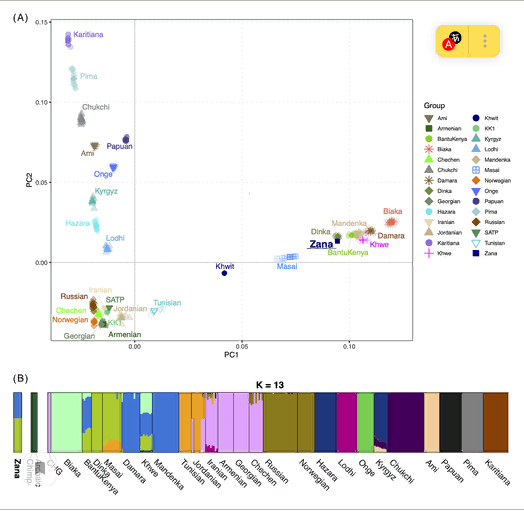
РИСУНОК 2
Откройте в Figure viewerPowerPoint
PCA и анализ примесей. А, PCA-анализ группы Zana, Khwit, HO современных популяций по всему миру и двух выборок CHG (KK1 и SATP). Четыре древних генома были спроецированы на современную вариацию. B, кластерный анализ на основе модели с использованием ПРИМЕСИ (K = 13) для оценки пропорций предков Заны. Всего для анализа было использовано 300 особей с 190 111 SNPs
Неконтролируемый кластерный анализ с использованием ПРИМЕСИ также явно отвергает любую гипотезу о том, что Зана была “нечеловеческого” происхождения, например, как это предлагается различными источниками.1, 2 Скорее, очевидно, что у нее была общая генетическая родословная с современными западно-и восточноафриканскими популяциями (рис. 2Б). Для дальнейшего изучения этого африканского происхождения мы провели дополнительный анализ PCA и примесей, основанный исключительно на африканских группах из группы HO (рисунок 3A и Рисунок S4Здесь снова Зана показывает родословные компоненты от восточных (например, Динка) и западных (например, йоруба)народов. Африканские группы, не имеющие значительного генетического вклада от популяций южной, северной и центральной Африки. Однако мы не смогли определить, была ли она (а) индивидуумом, происходящим от смешения между популяциями, похожими на динка и йоруба (компоненты пурпура и плинка на рис.3А), или (б) происходила исключительно из восточноафриканских групп, таких как Лухья и Ло.
РИСУНОК 3
Откройте в программе просмотра рисунковPowerPoint
Контролируемая/неконтролируемая примесь и анализы PCA (A, B и C, будут добавлены на график). A, Анализ кластеризации на основе модели с использованием ПРИМЕСИ (K = 7) с использованием ~примерно пяти человек из каждой из 48 африканских групп (n = 232) и 270 311 маркеров. B, пропорции генетического предка Заны с использованием K = 13 африканских потенциальных исходных популяций с использованием “контролируемого режима”, реализованного в ADMIXTURE. Результаты основаны на 300 репликах с различными начальными значениями. C, UMAP анализ Zana наряду с восточными, центральными и западноафриканскими группами
Чтобы более точно оценить долю предков Заны, мы провели анализ примесей в “контролируемом” режиме на основе 13 африканских популяций (рис. 3Б), представляющий большую часть разнообразия человеческих популяций в Африке. Несмотря на то, что результаты подтверждают, что Зана в основном восточная (~66%). Африканское происхождение, она также демонстрировала значительный уровень западноафриканского (~34%) генетического компонента. Для дальнейшей визуализации генетической связи Заны с африканскими популяциями мы применили дополнительное уменьшение размерности с помощью UMAP только с восточными, центральными и западноафриканскими группами, что показывает генетическую близость Заны с восточноафриканскими популяциями, такими как Ло и Лухья (рис. 3CЭто также подтверждается анализом максимального правдоподобия на основе TreeMix (рис. 4А ).
РИСУНОК 4
Откройте в программе просмотра рисунковPowerPoint
Филогенетическое дерево максимального правдоподобия и D-статистика. А, анализ TreeMix Zana и африканских популяций из группы HO. Это представляет собой дерево консенсуса, основанное на 100 репликах со случайными начальными значениями. Для анализа были использованы в общей сложности 98 особей из 22 популяций и 101 799 сайтов трансверсии (для уменьшения связанных с аДНК искажений в геноме Заны). Мы использовали уменьшенное число популяций для облегчения визуализации. Избыточный дрейфовый сигнал является артефактом псевдогаплоидной природы реконструированного генома Заны. B, 𝐷-статистика формы (Test_population, Zana; AltaiNea, Chimpanzee), предполагая, что в Zana нет значительного увеличения неандертальской примеси по сравнению с популяциями южнее Сахары (Z < 2), в то время как остальная часть мировой популяции демонстрирует хорошо известную неандертальскую примесь (Z > 2). Столбики представляют собой оценки ±2 SE
Чтобы формально проверить возможную интрогрессию неандертальца в геном Заны, мы провели 𝐷-статистику в виде (Test_population, Zana; AltaiNea, Chimpanzee) (рис. 4Б). Полученные результаты указывают на примесь неандертальцев только для популяций североафриканских и неафриканских популяций (хорошо известных из предыдущих исследований19, 38), что согласуется с генетической близостью Заны с субсахарскими группами, выявленными на основе кластерного анализа.Таким образом, наши результаты, основанные на общегеномном анализе, показывают, что геном Заны имел генетическое происхождение к югу от Сахары, что согласуется с результатами предыдущих (неопубликованных) краниометрических и мтДНК анализов. Это говорит о том, что ее присутствие в регионе, возможно, было связано с Османской империей работорговли в Стамбул, который был одним из основных центров для работорговли в регионе в 19 веке. Более того, большая часть восточноафриканских предков Заны согласуется с историческими записями, свидетельствующими о том, что большинство африканских рабов в Османской империи происходили из районов Великих Африканских озер и современного Судана.Современные сообщения и последующие рассказы о дикости Заны были, по крайней мере, частично основаны на некоторых ее необычных физических характеристиках, таких как отсутствие речи, умственная отсталость и длинные волосы, покрывающие все ее тело. Поскольку геномные данные явно отвергают все нечеловеческие гипотезы, мы предполагаем, что, если эти описания ее физических характеристик точны, у нее, возможно, было редкое генетическое заболевание человека, такое как врожденный генерализованный гипертрихоз: синдром с дисморфными чертами лица, умственной отсталостью и гипертрихоз.39
4 ВЫВОДЫ
Наши результаты доказывают, что неизвестной женщиной, похороненной на семейном кладбище Генаба, была сама Зана. В противоположность предположениям о том, что она могла быть женщиной-Алмастией, мы предоставляем окончательные данные по всему геному, чтобы положить конец рассказам о ней как о чем-либо, кроме человеческой женщины.Зана, вероятно, была восточноафриканского происхождения, хотя мы не можем исключить частичное западноафриканское происхождение. Мы предполагаем, что ее род мог появиться на территории современной Абхазии (Южный Кавказ) в результате работорговли, практиковавшейся между 16-19 веками Нашей эры Османской империей. Наконец, мы предполагаем, что это были просто ее незнакомые индивидуальные физические характеристики (такие как необычное поведение, физическая сила, высокий рост, отсутствие узнаваемой речи и гипертрихоз) и последующие слухи на протяжении поколений, которые подпитывали миф о нечеловеческом происхождении.
4.1 Записка по этике
После поимки в лесу Зана была лишена основных прав человека и обращалась с ней как с рабыней: ее держали в неволе, вероятно, заставляли вступать в сексуальные отношения с местными мужчинами и работали в условиях принудительного труда. После ее смерти рассказы о ее мифической фигуре привлекли нескольких ученых, чтобы раскопать ее историю, и кости ее сына были эксгумированы. Наше исследование направлено на то, чтобы раскрыть истинную человеческую природу Заны и даровать ей и ее потомкам то достоинство, которого они заслуживают.Все разрешения на раскопки в 1960-1970–е годы и на анализ аДНА были предоставлены соответствующими органами.
ПОДТВЕРЖДЕНИЯ
Мы благодарим Мирен Ираэту за помощь в лабораторных работах и Агнара Хельгасона за вдумчивое руководство. Игорь Бурцев благодарит Александра Машковцева и Бориса Поршнева за инициирование и организацию поисков и первого этапа раскопок; Николая Бурчака-Абрамовича, Владимира Орлкина, Юрия Воронова, Вадима Бжания, Леонида Яблонского и Владимира Никонова за поддержку и помощь в раскопках; Михаила Абдушелишвили, Александра Зубова, Галину Лебединскую, Марину Колодиеву, Маргариту Герасимову и Елизавету Веселовскую за помощь в антропологической работе.; Зеноби Чокуа, Нестор Думава (жители поселка Тхина, Абхазия), Ольга Кивилиди, Виктор Юшин (жители города Сухума) и местная администрация поселка Тхина-за помощь в раскопках; Таня Сабекия, Рая Сабекия (дочери Хвита), Родион Каджая, Ламара Каджая (потомки Заны), Анатолий Бганба (член семьи Хвита), Дмитрий Пиркулов, Олег Алиев, Михаил Шурдов и Григорий Вдовин-за помощь в раскопках. их помощь в восстановлении подробностей истории Заны инициировали и возглавили М. Томас П. Гилберт, Ашот Маргарян и Владимир Ямщиков. Ашот Маргарян, М. Томас П. Гилберт, Миккель-Хольгер С. Синдинг спроектировали это исследование. Миккель-Хольгер С. Синдинг, Ашот Маргарян, Владимир Ямщиков и Кристиан Каро подготовили данные. Ашот Маргарян, М. Томас П. Гилберт. а Владимир Ямщиков анализировал и/или интерпретировал полученные результаты. Рукопись написали Ашот Маргарян, М. Томас П. Гилберт, Владимир Ямщиков и Игорь Бурцев. Игорь Бурцев и Ашот Маргарян раскопали, кураторствовали, отбирали образцы и/или описывали анализируемые скелеты. Все авторы внесли свой вклад в окончательную интерпретацию данных.
АВТОРСКИЕ ВКЛАДЫ
Маргарян, Ашот: Концептуализация; Курация данных; Формальный анализ; Методология; Управление проектом; Надзор; Валидация; Визуализация; Написание-оригинальный проект; Написание-обзор и редактирование. Миккель-Хольгер Синдинг: Курация данных; методология; валидация; написание-обзор и редактирование. Кристиан Каро: Курация данных; исследование; методология; валидация; написание-обзор и редактирование. Владимир Ямщиков: Концептуализация; курация данных; методология; валидация; написание-рецензирование и редактирование. Игорь Бурцев: Концептуализация; исследование; методология; валидация; написание-рецензирование и редактирование. М. Томас Гилберт: Концептуализация; получение финансирования; расследование; управление проектом; надзор; написание-оригинальный проект; написание-рецензирование и редактирование.
КОНФЛИКТ ИНТЕРЕСОВ
Авторы не декларируют конкурирующих интересов.