Где-то в 2006 году была написана небольшая статья об эволюции генома-организма. Обычно, эволюция рассматривалась как приспособление организма к окружающей среде -адаптация. Я тогда рассмотрел вопрос о жизни самого информационного ядра- источника жизни и её сути. Дело в том, что в природе все сложные организмы содержат в своем геноме - следы мультипликации - копирования своих же частей. копируются кластеры, кусочки генов, гены , части хромосом, целые хромосомы и даже удваивается весь геном. Это невероятно гениальное изобретение Жизни, таким образом ,она легко УМНОЖАЕТ свой геномный материал, это ,впервые отметил японский ученый Сусуму Оно .
Уже ясно из известных примеров эволюционного развития разнообразия , что мультиплицирование – основной метод производства `материала` геномом для его прогрессирования . С другой стороны , такое , относительно гениально - простое и мощное решение проблемы материала для эволюции вызывает к жизни более драматическую картину происхождения и разнообразие путей эволюции.
Somewhere in 2006, a small article was written about the evolution of the genome-organism. Usually, evolution was considered as an adaptation of the body to the environment - adaptation. I then considered the question of the life of the information core itself - the source of life and its essence. The fact is that in nature all complex organisms contain in their genome - traces of animation - copies of their own parts. clusters, pieces of genes, genes, parts of chromosomes, whole chromosomes are copied and even the entire genome is doubled. This is an incredibly ingenious invention of Life, so it easily MULTIPLIES its genomic material, this, first noted by Japanese scientist Susumu Ono.
It is already clear from known examples of the evolutionary development of diversity that multiplication is the main method of producing 'material' by the genome for its progression. On the other hand, such a relatively ingenious - simple and powerful solution to the problem of material for evolution brings to life a more dramatic picture of the origin and variety of evolutionary paths.
Общая часть
Рассматривая сложноклеточные организмы , процессы мультиплицирования можно разбить на три основных типа :
1. Мультиплицирование одиночных кластеров - от единиц генов , до частей хромосомы.
2. Хромосом .
3. Целого генома.
Ясно , что процессы сопровождающие такие созидающие `катаклизмы` в них будут отличаться массивностью перекрытия активных частей генома . При удвоении кластера будет наблюдаться перекрытие его активных частей и, соответствующее напряжение в геноме . Существующие механизмы регуляции в геноме , в состоянии ограничить их общую активность . Вплоть до ингибирования одного из них . Такое ингибирование сделает возможным быструю мутацию неактивного компонента и его переход в другую `плоскость` работы . Ясно , что при удвоении хромосомы , подобное явление уже будет массовым и напряжение в геноме будет намного большим. Здесь мы рассматриваем выживающие после подобных явлений особи . И две хромосомы через некоторое время уже будут сильно отличаться от `материнской` рис 1..
General part
Considering complex cell organisms, multiplication processes can be divided into three main types:
1. Multiplication of single clusters - from gene units to parts of the chromosome.
2. Chromosome.
3. Whole genome.
It is clear that the processes accompanying such creative 'cataclysms' in them will differ in the massiveness of overlapping the active parts of the genome. When the cluster is doubled, the overlap of its active parts and the corresponding voltage in the genome will be observed. Existing mechanisms of regulation in the genome, able to limit their overall activity. Up to inhibition of one of them. Such inhibition will allow rapid mutation of the inactive component and its transition to another 'plane' of operation. It is clear that when the chromosome doubles, such a phenomenon will already be massive and the tension in the genome will be much greater. Here we consider individuals surviving after such phenomena. And after a while, the two chromosomes will already be very different from the 'maternal' of Fig. 1..
РИС 1
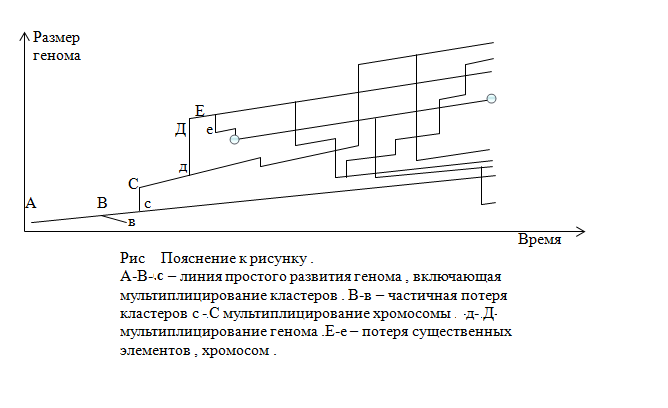
Материнская хр-ма – 1 , 2 , 3 , 4 , 5 , 6 , 7 , 8 , 9 , 10 , 11 , 12 , 13 ,14 ...
І (от удв.) хр-ма - 1 , 2 , 3 , 0 , 5 , 6 , 0 , 0 , 0 , 0 , 11 , 0 , 13 , 0 ...
ІІ (от удв.) хр-ма - 0 , 0 , 0 , 4 , 0 , 6 , 7 , 8 , 9 , 10 , 0 , 12 , 0 ,14 ..
пояснение к Рис 1 Показаны гипотетичная структура
Хромосом – Материнской и двух удвоенных І , ІІ .
Цифры 1,2,3,4,... показывают активные компоненты
материнской хромосомы .
Ясно , что вместо `0` в определенный момент появляется новый активный компонент , но этот компонент уже будет `сгенерирован` только в данной хромосоме . Таким образом , через определенное время , мы уже с трудом можем принять , что две хромосомы связаны дублированием одной материнской .
Примерно тоже произойдет и при удвоении целого генома , только процесс будет глобальней .
Возникает интересное следствие из этого представления . Уравновешение активных компонент генома может идти по двум основным направлениям : `ускоренная` мутация и вывод из сферы действия другого двойника и просто `потеря` - исключение его из генома . Ясно , что второй процесс – теряет накопленное , но , он происходит и быстро решает внутренние напряжение . Тогда , картина эволюции , уже выглядит достаточно сложнее . Становится очевидным , что геном может идти несколькими путями :
1. Наращивая размер и отрабатывая механизм `погашения` напряжений.- Медленное движение вверх в иерархии развития.
2. Наращивая , уплотняя информацию и теряя размер для `погашения` напряжений.- Революционные скачки в усложнении и движении вверх в иерархии развития .
3. Наращивая размер и теряя его – падая на более нижний уровень в иерархии развития
и /или в размере генома .
FIG 1
Maternal chrome (MC) - 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14...
І (from doubling) MC - 1, 2, 3, 0, 5, 6, 0, 0, 0, 0, 11, 0, 13, 0...
ІІ (from doubling) MC - 0, 0, 0, 4, 0, 6, 7, 8, 9, 10, 0, 12, 0, 14..
explanation of Figure 1 A hypothetical structure is shown
Chromosomes - Mother and two doubled I, II.
Figures 1,2,3,4,... show active components
maternal chromosome.
It is clear that instead of '0', a new active component appears at a certain point, but this component will already be' generated'only on this chromosome. Thus, after a certain time, we can hardly accept that two chromosomes are connected by duplication of one maternal.
Approximately the same will happen with the doubling of the entire genome, only the process will be more global.
An interesting consequence arises from this idea. Balancing the active components of the genome can go in two main directions: 'accelerated' mutation and withdrawal from the scope of another twin and simply 'loss' - excluding it from the genome. It is clear that the second process - loses the accumulated, but, it occurs and quickly solves internal tension. Then, the picture of evolution already looks rather more complicated. It becomes apparent that the genome can go in several ways:
1. Increasing the size and practicing the mechanism of 'repayment' of tensions. - Slow upward movement in the hierarchy of development.
2. Building up, compacting information and losing size to 'pay off' stresses. - Revolutionary leaps in complication and upward movement in the hierarchy of development.
3. Growing size and losing it - falling to a lower level in the development hierarchy
and/or genome size.
Рис 2 FIG 2
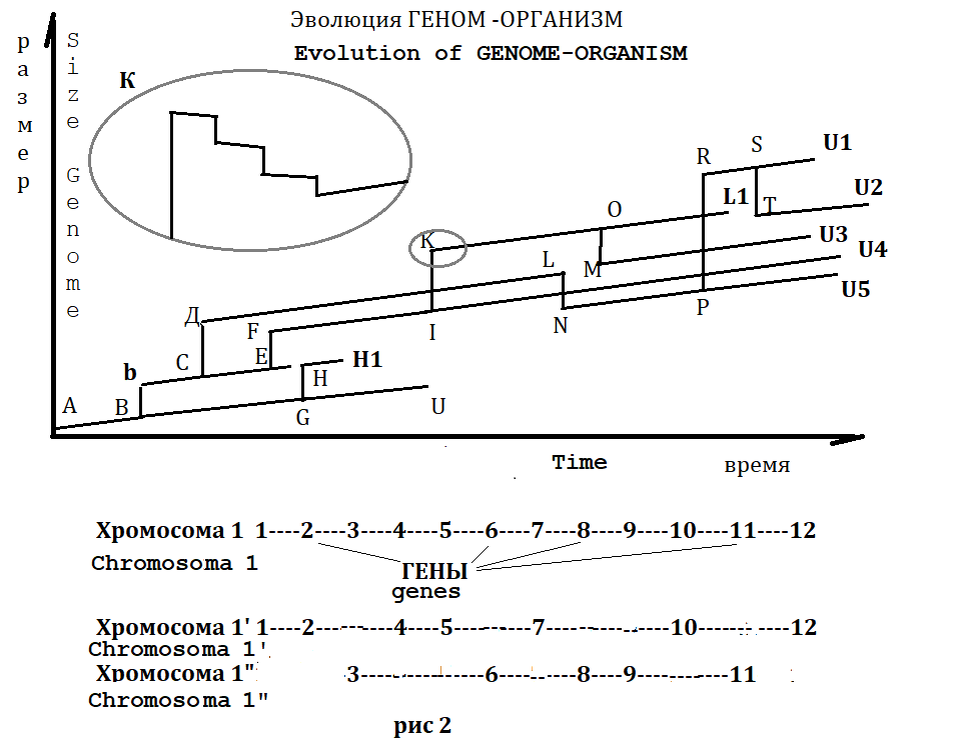
Пример регрессивного движения Генома :
http://genomebiology.com/2007/8/4/R59/abstract
Недавно, целые последовательности генома стали доступными для двух cnidarians, Hydra magnipapillata и Nematostella vectensis, и большие наборы данных EST для них и для коралла Acropora millepora. Результаты позволили лучше понимать основание врожденной неприкосновенности в cnidarians, мы просмотрели доступное EST и геномные ресурсы для некоторых из ключевых компонентов позвоночного врожденного свободного репертуара, фокусируясь на тропах дополнения и Toll/TLR. Каноническая тропа Toll/TLR присутствует в представителях основного cnidarian Класса, Anthozoa, но ни классик Toll/TLR-рецептор, ни обычный NF-kb не мог быть идентифицирован в морском членистоногом животном Hydra. Кроме того, обнаружение дополнения, C3 и несколько белков MAC/PF предполагают, что prototypic тропа исполнительного элемента дополнения может существовать в морских членистоногих животных, но не в hydrozoans. Вместе с данными для нескольких других генных семей, это подразумевает, что Hydra, возможно, подвергся существенной вторичной генной потере в течение развития. Такие потери не ограничены Hydra, однако, и по крайней мере один ген MAC/PF, кажется, потерян от Nematostella. Рассмотрение этих образцов генного распределения подчеркивает вероятное значение генной потери в течение развития животных, указывая древнее происхождение для многих компонентов позвоночной врожденной иммунной системы.
An example of Genome regression motion:
http://genomebiology.com/2007/8/4/R59/abstract
Recently, whole genome sequences have become available for two cnidarians, Hydra magnipapillata and Nematostella vectensis, and large EST datasets for them and for the coral Acropora millepora. The results provided a better understanding of the basis of innate immunity in cnidarians, we looked at available EST and genomic resources for some of the key components of the vertebrate innate free repertoire, focusing on complement and Toll/TLR trails. The Toll/TLR canonical pathway is present in representatives of the major cnidarian Class, Anthozoa, but neither the Toll/TLR receptor classic nor the common NF-kb could be identified in the marine arthropod animal Hydra. In addition, the discovery of complement, C3, and several MAC/PF proteins suggest that the prototypic pathway of the complement executive element may exist in marine arthropods, but not in hydrozoans. Together with data for several other gene families, this implies that Hydra may have undergone substantial secondary gene loss during development. Such losses are not limited to Hydra, however, and at least one MAC/PF gene seems to be lost from Nematostella. Consideration of these gene distribution patterns highlights the likely significance of gene loss during animal development, indicating ancient origins for many components of the vertebral innate immune system.
РИС 3 (из работы (2) ) FiG 3 (From paper (2))
Как видим рисунок 3 идеально совпал с предложенной моделью эволюции. Только авторы не определяли Размер геномов в этом эксперименте. Но, очевидно, что мы увидим примерно предложенный график 1.Только отличие будет в том, что делеции, будут следовать сразу за массивной мутацией, что просто очевидно.
As you can see, Figure 3 perfectly coincided with the proposed model of evolution. Only the authors did not determine the size of the genomes in this experiment. But, obviously, we will see roughly the proposed graph 1. Only the difference will be that deletions will follow immediately after the massive mutation, which is simply obvious. FIG 4
РИС 4 FIG 4
Таким образом. сами свойства ДНК, и методы решения сложных проблем развития во времени породили эту эволюцию,которая сцеплена с возможностями сохранения и переработки в эволюционном развитии -геномного сырья.
Эволюция оказалась сложнее и мощнее чем представлялось нам раньше в Дарвинистких моделях развития.
Thus. the very properties of DNA, and methods for solving complex problems of development over time, gave rise to this evolution, which is intertwined with the possibilities of conservation and processing in the evolutionary development of genomic raw materials.
Evolution turned out to be more complex and powerful than previously imagined in Darwinian development models.
Литература Literature
1 Susumu Ohno “Evolution by gen dublication”
2 "The Dynamics of Molecular Evolution Over 60,000 Generations"
Benjamin H. Good,1,2,3,4,5,* Michael J. McDonald,1,2,6,* Jeffrey E. Barrick,7,8 Richard E. Lenski,8,9 and Michael M. Desai1,2,
Nature. 2017 Nov 2; 551(7678): 45–50.
Published online 2017 Oct 18. doi: 10.1038/nature24287