Продолжение
Хорошо известно, что допамин генерирует супероксидные анионы и хинон допамина путем его автоокисления ( Klegeris et al., 1995). Полученный хинон дофамина становится ковалентно конъюгированным с сульфгидрильной группой цистеиновых остатков белков (Kuhn et al., 1999; Whitehead et al., 2001 ), приводя преимущественно к образованию хинопротеинов ( Graham, 1978 ; Fornstedt et al., 1986). Антиоксидант NAC, как известно, ингибирует окисление дофамина до хинона дофамина, очищает свободные радикалы, такие как гидроксильные радикалы, и ингибирует образование хинопротеинов путем связывания с хиноном дофамина ( Pedrosa and Soares-da-Silva, 2002 ; Khan et al., 2003; Izumi et al., 2005). Поэтому для выявления участия супероксидных анионов и дофаминхинона в эффектах дофамина нами было изучено влияние NAC на дофамин-индуцированное ослабление ЛПС-индуцированной экспрессии цитокинов. NAC (10 мм) был эффективен в обращении вспять этого ослабления ЛПС-индуцированной мРНК и белковой экспрессии цитокинов, вызванных 30 мкм дофамина ( Рис.2). 3 А-Ф). С другой стороны, предварительная обработка клеток Н2О2 (10-100 мкм) не была эффективной, как и комбинация ксантиноксидазы (1-10 МЕ/мл ) и гипоксантина (50 мкм), который генерирует супероксидные анионы ( Рис.2). 3 G-I).
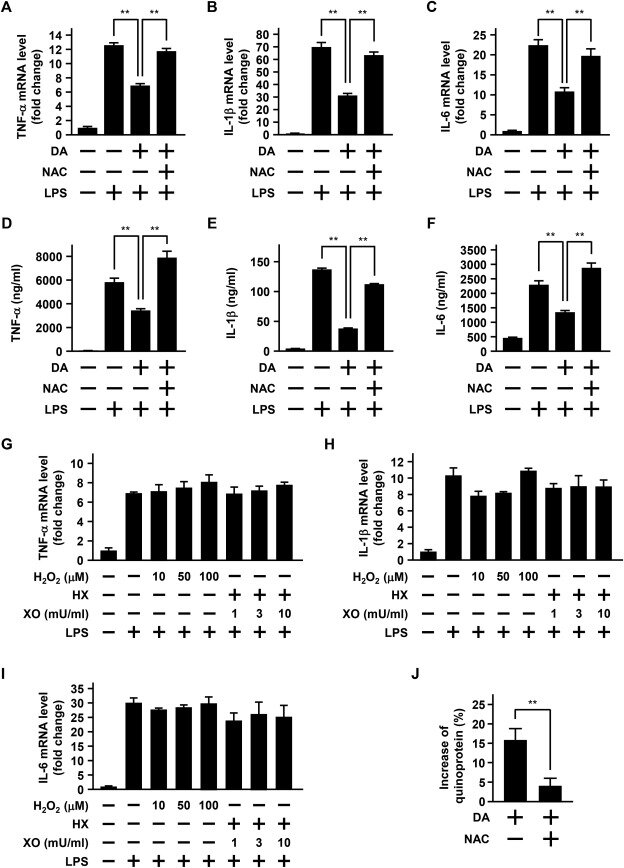
Далее мы оценивали образование хинопротеинов после обработки дофамином клеток БВ-2. Обработка клеток 30 мкм дофамином в течение 24 ч повышала уровень хинопротеинов в клетках, и это повышение ингибировалось 10 мм NAC ( Рис.2). 3J).
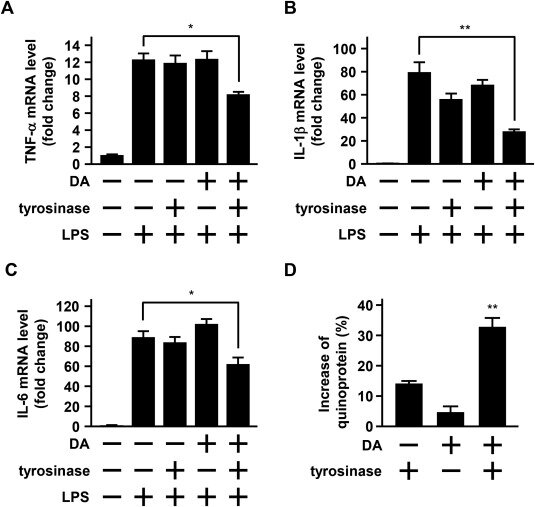
Предварительная обработка 30 мкм дофамином в течение 1 ч не смогла ослабить индуцированную ЛПС ( 10 мкг/мл) экспрессию мРНК цитокинов (Рис.2). 4 А–С). С другой стороны, в присутствии 300 Ед/мл тирозиназы, катализирующей окисление дофамина до хинона дофамина, предварительная обработка 30 мкм дофамина существенно ослабляла экспрессию ЛПС-индуцированной мРНК этих цитокинов. В присутствии 300 Ед/мл тирозиназы обработка 30 мкм дофамина в течение 1 ч значительно повышала уровень хинопротеина ( Рис.2). 4D), предполагая, что дофамин ослаблял индуцированную ЛПС экспрессию мРНК этих цитокинов через образование хинона дофамина в клетках BV-2.