ГЕНЕЗИС ЧЕЛОВЕКА
(Очерк популяционной генетики)
2014г.
Генезис человека
(очерк популяционной генетики)
Потребность в написании этой статьи возникла случайно. Это произошло после того, как я стал дедушкой. 16 января 1999 года родился мой первый внук Артем. И вдруг я ощутил себя «предком», - одним из звеньев цепи, протянувшейся через века и времена. Раньше я не задумывался о том, что эта живая цепь тянется непрерывно – от самых первых одноклеточных организмов до меня лично и далее к моим потомкам. Сейчас у меня уже пять внуков. Для своих внуков и будущих пра-пра-внуков я – предок. И у меня тоже были предки. А кто они? Что я знаю о своих предках? И вообще, сколько моих предков было?
Мы знаем чаще всего только своих ближайших родственников в двух – трех поколениях. Очень немногие люди могут проследить свою генеалогическую линию на пять поколений, и только единицы – на десять поколений назад. А ведь десять поколений – всего лишь 300 лет истории! Начнем с, казалось бы, простого вопроса: сколько всего у меня было предков?
В контексте биологической эволюции видимо следует проследить живую цепь по нисходящей линии от конкретного человека до первобытных людей, обезьян, динозавров, простейших одноклеточных организмов. Очевидно, что в биологическом смысле самыми первичными предками могут считаться только многоклеточные организмы, имеющие разделение полов. Только для них можно говорить о передаче наследственной информации по прямой генетической линии, то есть о детерминистическом состоянии генов каждого организма. Предшествующие организмы, не разделенные по полу, имели «несеперабельное» состояние генетической информации, которое можно рассматривать как суперпозицию (одновременное наложение) всех генов определенной группы организмов. А одноклеточные организмы вообще следует считать нашими общими предками в биологическом смысле, вплоть до самой первой возникшей из органической материи структуры ДНК, способной к самовоспроизводству. Однако такое глубокое исследование мы полагаем нецелесообразным. Достаточно ограничиться временем появления приматов вида «Homo sapiens» и выхода его из первоначального ареала обитания в Африке, то есть примерно 70-80 тысяч лет назад.
Таким образом, наше исследование будет касаться только биологических (генетических) предков человека, которых мы в дальнейшем будем называть «прямыми предками». Поэтому название «генезис человека» точно отражает задачи нашего исследования.
В дальнейшем под «генезисом человека» будем подразумевать происхождение конкретного индивидуума, человеческой особи, понимаемого нами как процесс передачи генетической информации между поколениями.
Для целей нашего исследования необходимо определить некоторые исходные положения, чтобы не запутаться в многообразии вариантов ответов на поставленные вопросы.
Примем первый постулат, который можно назвать «ЭГО-логическим»:
Если «Я» существую, значит существовали ВСЕ мои прямые предки.
Этот постулат является необходимым и достаточным условием моего существования. Для того чтобы в конце цепи появился один человек, необходимо чтобы в цепи были в наличии все звенья – его прямые предки!
Второй постулат вытекает из первого:
В каждом поколении должны существовать ОДНОВРЕМЕННО мои прямые предки разного пола вступающие в брак.
Этот постулат основывается на биологических особенностях разделения полов – чтобы родился новый организм, одновременно должны быть в наличии отец и мать. Понятие одновременности разумеется относительное, только применительно к моменту зарождения потомства. Одновременность следует понимать как период времени жизни одного поколения в несколько лет, что несущественно для общей продолжительности анализируемых периодов времени.
Одновременное наличие в поколении двух прямых предков разного пола и половые связи между ними, в результате которых появляется потомство, мы в дальнейшем будем называть «браком».
Время воспроизводства одного поколения прямых предков можно принять 25 лет. Конечно, есть примеры вступления в «брак» человеческих особей в возрасте достижения половой зрелости – 12-13 лет, и рождения детей в возрасте 45-60 лет, но наиболее типичным для «брака» является возраст 25 лет. Это относится и ко времени рождения второго, третьего и более младшего ребенка в одном «браке». В ранние периоды развития человечества продолжительность жизни одного индивидуума (предка) была меньше – до 25 лет и даже до 18 лет, однако с учетом неуклонного увеличения общей продолжительности жизни и все более позднего возраста вступления в «брак» мы полагаем вполне корректным принять среднее время жизни (воспроизводства) одного поколения – 25 лет. Соответственно за 100 лет сменяется 4 поколения.
Так сколько у меня было прямых (биологических) предков?
Вопрос о количестве моих предков не такой простой, как кажется на первый взгляд.
Первый поверхностный ответ действительно прост: в каждом предыдущем поколении количество предков должно удваиваться. Ведь для рождения одного потомка необходимо наличие двух особей предков - мужчины и женщины.
Если принять что количество прямых предков в одном поколении «N», число поколений (номер поколения при обратном отсчете) - «n», то:
N=2n (1)
То есть, если принять мое «я» за нулевое поколение, то при обратном отсчете (регрессии) в первом поколении будет два предка, во втором поколении – четыре, в третьем – восемь, в четвертом – шестнадцать, в пятом – тридцать два и так далее.
Для наглядности расчеты можно проиллюстрировать следующей схемой:
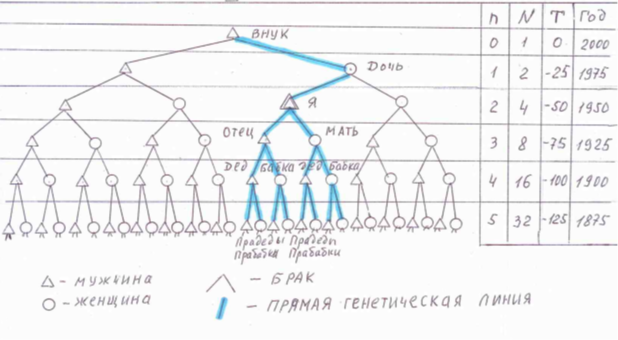
На схеме можно проследить прямые генетические связи между поколениями, соответственно наглядно видно, что число необходимых предков удваивается в каждом предыдущем поколении.
В соответствии с сформулированными первым и вторым постулатами можно сказать, что к примеру, 100 лет назад на Земле одновременно проживали 16 моих прямых предков, а 200 лет назад количество одновременно проживающих на Земле моих прямых предков должно быть 256 человек.
Однако, простой расчет, например для 40-го поколения (примерно на 1000 лет назад) показывает, что количество одновременно проживающих на Земле моих прямых предков должно составлять 1 099 510 839 296 человек (один триллион девяносто девять миллиардов пятьсот десять миллионов восемьсот тридцать девять тысяч двести девяносто шесть человек!).
Согласно научным данным численность населения всей Земли составляла:
в 4000-м году до н.э. – 7 млн. человек;
в 1000-м году до н.э. – 50 млн. человек;
в 1-м году н.э. – 290 млн. человек;
в 1000-м году н.э. – 380 млн. человек;
в 1650-м году – 545 млн. человек;
в 1800-м году – 907 млн. человек;
в 1900-м году – 1 610 млн. человек;
в 1950-м году - 2 509 млн. человек;
в 1987-м году – 5 026 млн. человек;
в 2000-м году – 6 070 млн. человек;
в 2014-м году – 7 200 млн. человек.
Таким образом, расчетная численность моих предков живших тысячу лет назад и совершенно необходимых для моего появления на свет, на 4 порядка (тридцать тысяч раз!) больше чем все население Земли в это время!
Эту несуразность нельзя объяснить иначе, как путем признания факта, что закон удвоения численности предков не соответствует действительности.
Какой же фактор ограничивает рост численности моих предков в одном поколении? И каков математический закон позволяющий рассчитать реальное число прямых предков в обратной регрессии?
Мы должны признать, что такие естественные ограничения роста численности популяции человечества как ограниченность пищевой и ресурсной базы, изменения климата, болезни, не могут служить таким фактором. В самом деле, уменьшение численности и даже вымирание части популяции человеческого общества не может оказать влияние на число моих предков в силу первого постулата. Те, которые вымерли, не произведя потомства, не были моими предками!
В силу второго постулата моими прямыми предками в каждом поколении могли быть только одновременно проживавшие люди имевшие потомство.
Единственным приемлемым объяснением указанного феномена может служить предположение, что в одном поколении в «брак» между собой вступали одновременно несколько особей, находящихся в «кровном» (генетическом) родстве. Такие «браки» мы называем «перекрестными».
Следует отметить, что речь не идет о полигамии или моногамии. Полигамия – «брак» между одним мужчиной и несколькими женщинами, или между одной женщиной и несколькими мужчинами. Моногамия – «брак» между одним мужчиной и одной женщиной. Мы полагаем, что необходимо четко определить, что только в случае «перекрестных браков» имеет место сужение генетической базы воспроизводства потомства. В самом деле, только «браки» между близкими родственниками (мужчиной и женщиной) приводит к уменьшению общего числа особей, участвующих в передаче генетической информации. Например, полигамный «брак» одного мужчины с тремя женщинами, имеющими разные генетические линии, не приводит ни к увеличению, ни к уменьшению количества необходимых предков в предыдущих поколениях по сравнению с законом удвоения. И наоборот, всего лишь один моногамный «перекрестный брак» между близкими «кровными» родственниками, например между мужчиной и женщиной являющимися братом и сестрой, приводит к уменьшению числа предков в предыдущих поколениях, вплоть до стагнации.
Сами по себе «перекрестные браки» (полигамные и моногамные) между родственниками не могут быть приняты в качестве причины ограничения роста численности предков, без введения еще одного, третьего постулата:
В каждом поколении «перекрестные браки» между близкими кровными родственниками запрещены (между отцом, матерью и детьми, между братьями и сестрами, между дедом, бабушкой и внуками).
Обозначая через «p» число «перекрестных браков» в одном поколении с номером «n», математически этот постулат можно выразить следующим образом: pn ≤ n – 2 , где n>3 (2)
Соответственно необходимое количество предков «N» в поколении «n» будет выражаться формулой: N = 2n - p (3)
Для наглядности изложения теории генезиса иллюстрацией может служить следующая схема:
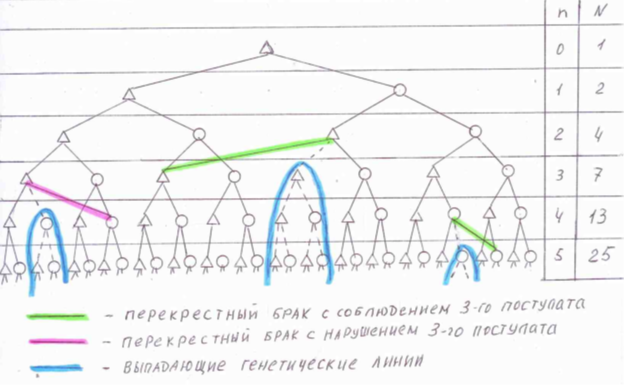
Из этой схемы становится понятным, почему мы назвали такие браки «перекрестными».
Если предположить, что в каждом поколении «n» допускается всего один «перекрестный брак» между родственниками, число необходимых прямых предков в этом поколении будет равно N = 2n - 1. Для рассмотренного выше примера (100 лет назад с числом поколений 4), количество предков будет равно 15 человек, а не 16. Однако с увеличением числа перекрестных браков в каждом поколении этот показатель быстро уменьшается, что приводит к существенному сужению генезисной базы человека. Одна из генетических линий при «перекрестном браке» обрывается, соответственно в каждом предыдущем (нисходящем) поколении число предков по этой прерванной линии будет увеличиваться в два раза. Например, если в поколении «n» будет один перекрестный брак, число предков в этом поколении уменьшится на 1, в поколении «n+1» на 2, в поколении «n+2» на 4, в поколении «n+3» на 8, и так далее. Если допустить что в каждом поколении будет только один «перекрестный брак», то число «оборванных» генетических линий будет увеличиваться в геометрической прогрессии.
«Браки» между родственниками не такая уж редкость даже в наши дни. Более того, в условиях небольших по численности и изолированных групп особей, неизбежно все члены группы через определенное время становятся родственниками. Несомненно, что в более ранние эпохи развития цивилизации браки между родственниками были более широко распространены в силу затруднений в коммуникациях и отсутствия обмена генетическим материалом.
Однако, допустимыми являются только браки между двоюродными родственниками и более дальними «по крови». Для введения этого постулата оправданием может служить многовековой интуитивный опыт человеческой цивилизации, говорящий о том, что браки между близкими кровными родственниками быстро приводят к вырождению потомков вследствие накопления мутаций и дефектов генов. Это требование присутствует практически во всех цивилизациях. Даже если нет императивного законодательного или морального запрета, все равно в обычаях и традициях всех народов и племен этот запрет присутствует как «неодобрение». Более того, почти у всех народов весьма почетным делом было похищение женщин и женитьба на представителях другого рода, племени, деревни. Взять жену или выйти замуж следовало как можно дальше от «своих». В этом проявлялись интуитивные подсознательные запреты «кровосмешения» и потребности «влить свежую кровь», которые мы изложили в виде третьего постулата.
Третий постулат определяет количество и время цикла реализации неповторяющихся комбинаций особей в «перекрестных браках» более дальних «по крови», чем двоюродные.
Разумеется, этот постулат может использоваться в пределах времени существования человеческой цивилизации – примерно до 20-30 тысяч лет назад. В более ранние периоды и в малочисленных популяциях отношения полов могли быть близкими к животным биологическим, однако как мы увидим в дальнейшем, этот постулат применим даже для ограниченных популяций человеческих особей.
Комментируя указанную формулу (3) следует отметить, что в ней не учитывается влияние количества «перекрестных браков» в предыдущих поколениях «mn» и количество повторяющихся «перекрестных браков» «Sn,m». Это очень существенно влияет на расчет числа «N», но как минимум не увеличивает это число, а уменьшает. Кроме того, для учета влияния «перекрестных браков» на количество предков следует учитывать общую численность изолированной популяции. Влияние всех факторов носит, в общем, случайный характер, подчиняющийся законам вероятностей. Математически вывести формулу для расчета числа предков можно используя методы статистической генетики и теории вероятности. Однако, выведение общей формулы для расчета числа предков в любом поколении с учетом всех перекрестных браков является сложной математической задачей и в данной работе не рассматривается.
Для регрессного расчета количества предков в каждом поколении при условии, что в каждом поколении имеет место только один «перекрестный брак» формула приобретает вид:
Nn = (2n – 2n-2) + 1, или Nn = 3*2n-2 + 1 (4)
Пример расчета для вышеуказанных условий приведен в таблице 1:
При наличии более чем одного «перекрестного брака» в одном поколении количество прямых предков в предыдущих поколениях стремительно уменьшается. Возможны различные варианты развития «дерева» популяции, в том числе и стабилизация численности на определенном уровне.
Наиболее интересен случай, когда в каждом поколении реализуется максимально возможное количество «перекрестных браков» при условии соблюдения третьего постулата. Это возможно даже при относительно небольшом числе особей в поколении, ранее не связанных общей генетической линией.
Пример расчета для популяции численностью 8 особей (4 мужчины и 4 женщины) приведен в таблице 2:
Расчет можно проиллюстрировать следующей схемой:
Разумеется, указанный пример развития популяции с учетом «перекрестных браков» не является единственным. Возможны различные неповторяющиеся комбинации «браков» между особями.
Из приведенных расчетов и схемы следует, что численность прямых предков в предельном случае может не увеличиваться, а стабилизироваться на значении, достаточном для выполнения третьего постулата в течение нескольких поколений. Сужение базы генезиса в этом случае вызвано исключительно фактором «перекрестных браков», что подтверждает выдвинутую нами теорию генезиса человека.
Также интересен случай, когда изначально очень небольшое количество предков вступали в «перекрестные браки» с нарушением третьего постулата. Речь идет о размножении популяции, по каким либо причинам получившей достаточную пищевую базу и благоприятные условия проживания при расселении на новых территориях.
Можно представить небольшую группу людей переплывших море или перешедших через горы и оказавшихся в благоприятных условиях, но изолированных от других групп. Количество особей в каждом поколении при этом не ограничено, но количество прямых предков все равно остается ограниченным. Например, если проследить генетическую линию в четырех поколениях прямых предков в популяции, возникшей первоначально из одной пары (мужчины и женщины) и принимая, что у каждой пары особей было 4 потомка, то при «n = 0» «N=1», получим, что число предков в поколении «n = 1» составляет 2, в поколении «n = 2» - максимум 4, в поколении «n = 3» опять 4, и в поколении «n = 4» - 2!
Общая численность такой группы может стремительно увеличиваться, так как количество потомков в одном «браке» может достигать 10-15 и более. Ограничением здесь является чисто биологическая способность женского организма к воспроизводству (возможное количество детей в детородном возрасте). Конечно, все они будут родственниками, в том числе близкими «кровными» родственниками, иметь общих родителей или бабушек, дедушек. Важно отметить, что число прямых предков при «перекрестных браках» между близкими «кровными» родственниками тоже ограничено N = 2n-1 – p (3).
Мы хотим сказать, что несмотря на рост общей численности популяции в геометрической прогрессии K = 2n, число прямых биологических предков в одном поколении будет ограничено общим количеством особей в популяции как максимум N= 2n-3. При определенных условиях, при n> 8, число прямых предков вообще может быть не зависимым от «К». Мы покажем ниже, что число предков в одном поколении может не превышать 256 особей.
Попытаемся проанализировать некоторые выводы, вытекающие из изложенной теории генезиса человека.
Первый вывод состоит в том, что каков бы ни был закон расчета численности предков в одном поколении, он содержит показательную функцию (2n) с различными коэффициентами регресса. Это означает неизбежное увеличение числа N, пока не наступит полное исчерпание генезисной базы популяции. Мы хотим сказать, что количество особей в каком-то регрессном поколении будет равно общему количеству проживающих людей на Земле. Все люди – родственники! Этот вывод подтверждается данными науки антропологии, свидетельствующими о том, что все человечество произошло от одной малочисленной группы особей вида «Homo sapiens», первоначально проживавшей в маленьком ареале обитания в Африке около 80 тысяч лет назад.
Второе следствие состоит в том, что возможно ограничение генезисной базы популяции, проживающей в изоляции от других популяций в течение достаточно больших периодов времени без нарушения третьего постулата.
Поясним это на примере. Предположим, что в 1600 году н.э. число особей в изолированной популяции, например, в деревне далеко в горах, составляет 16 человек (8 мужчин и 8 женщин) и ограничено возможностями пищевой базы среды обитания. Тогда популяция может сохранять свою численность без «перекрестных браков» между близкими родственниками (третий постулат) в течение 16 поколений. С учетом принятой нами продолжительности цикла воспроизводства одного поколения 25 лет, время существования этой группы с неповторяющимися комбинациями «браков» между особями составит 400 лет (!). То есть, при наличии всего лишь 16 индивидуумов такая группа благополучно дожила бы до наших дней, сохранив свой первоначальный генетический фонд без вырождения.
Продолжая анализ с учетом социально – культурологических условий общества за последние 200 лет, можно предложить следующую модель регрессного генезиса человека.
За нулевую точку отсчета примем 2000 год н.э., и примем, что в течение 8-ми поколений (200 лет) браки между предками заключались только из разных генетических линий, т.е. не между родственниками. Число необходимых предков в 1800 году н.э. составит 256 человек. Предположим, что по каким-то причинам до 1800 года н.э. эта группа проживала в изоляции. Допустим, что браки внутри группы в этот более ранний период времени заключались с учетом ограничений, накладываемых третьим постулатом. Тогда получается, что минимальное число необходимых прямых предков составляет всего 256 человек в каждом из предыдущих 256 поколений в течение 6400 лет (!), то – есть, начиная с доисторических времён! Такое резкое уменьшение генезисной базы является вполне допустимым с биологической точки зрения. Происходит стабилизация численности предков в течение длительного времени.
Это положение подтверждается историческими, археологическими и антропологическими данными. Человеческая популяция, небольшая по численности, начавшая движение около 80 тысяч лет назад, в процессе расселения и освоения новых территорий неизбежно не только росла количественно, но и разбивалась на отдельные компактные группы, длительное время не контактировавшие между собой (народы, племена, рода). Это означает, что реальное количество прямых предков не может быть очень большим. Поскольку расчеты имеют вероятностный характер и достаточно сложны, мы даем экспертную оценку, что максимальное число прямых предков может не превышать численность первоначальной группы «Homo sapiens» - 4-5 тысяч особей.
Третье следствие из теории генезиса человека заключается в том, что периоды стабилизации численности предков имеют важнейшее значение для эволюции вида.
Этот «застой» на самом деле означает концентрацию генетического материала. При обычном расширенном воспроизводстве поколений неизбежно происходит смешивание и рассеяние генетической информации. При расширенном воспроизводстве этот процесс совершенно необходим для расширения генетической базы вида и возникновения новых мутаций. Однако он увеличивает время возникновения и закрепления полезных мутаций, чаще всего большинство мутаций подавляется. Вид развивается экстенсивно, и задачей естественного отбора в это время является сохранение генома и передача его без изменений следующим поколениям. Во время цикла стабилизации численности изолированной группы вида, генетические изменения не рассеиваются, а накапливаются. Вид начинает развиваться интенсивно, вовлекая в процесс естественного отбора приобретенные и сохраненные ранее гены. Они могут дать новое качество, новый полезный признак, закрепить его в генах целой группы особей за сравнительно короткое время. Получившие новое качество члены группы становятся «пассионарными», т.е. движущей силой эволюции.
Здесь можно высказать те же соображения математического – логического толка, основанные на теории управления в кибернетике. Если численность популяции слишком мала (меньше восьми особей), уже через несколько поколений неизбежно появление «перекрестных браков» между близкими родственниками, т.е. нарушение третьего постулата. Как видно из практики, потомство близких родственников слишком быстро накапливает мутации, зачастую имеющие негативное влияние на устойчивость и жизнеспособность потомства. Это аналогично наличию положительной обратной связи в кибернетической системе управления каким-либо объектом: входной сигнал складывается с выходным, усиливается и входит в резонанс, с разрушительными для объекта последствиями. Система управления при этом уже не передает никаких сигналов, процесс передачи информации прекращается. На практике в генетической биологии это означает невозможность для особи уже в третьем – четвертом поколении близкородственных браков произвести потомство (через 50-100 лет).
Если количество особей в замкнутой группе относительно велико (несколько десятков или сотен), процесс накопления и передачи генетической информации носит более плавный характер. При этом негативные эффекты аналогичные резонансу в системе управления, гасятся в массе другой генетической информации. Единичные особи, получившие чрезмерную сумму одинаковых генов, гибнут или не дают потомства. Усиливаются и передаются только те информационные сигналы (гены), которые одновременно сохраняются в генотипах большого числа группы особей, так как оказываются полезными для большого числа их носителей. Процесс накопления положительных генов по нашей экспертной оценке может занимать около 10-15 поколений, или по нашей временной шкале – 250 – 400 лет.
Обобщая выводы теории генезиса следует обратить внимание на следующее.
Как мы показали выше, количества прямых предков в 256 человек оказывается вполне достаточным для анализа генетического дерева в исторический период европейско-азиатской цивилизации. Необходимо также учитывать относительную изолированность отдельных популяций, например, индейцев Америки или аборигенов Австралии, которые во всяком случае не могли быть моими прямыми предками в течение последних 6400 лет.
Если продолжить идеализированную теоретическую экстраполяцию процесса генезиса дальше во мглу веков и тысячелетий, получим следующие результаты:
- в период с 6400 лет до 102 тысяч лет минимально необходимое и достаточное число моих предков составляет 4096 особей.
- в период от 100 тысяч лет до 1,6 миллиона лет число предков может составлять не более 65,5 тысяч особей.
Дальше нет смысла теоретизировать.
Это поразительно! Оказывается, чтобы я появился на свет, необходимо и достаточно, чтобы 80-100 тысяч лет назад в Африке жила группа первобытных людей численностью всего 4-5 тысяч особей! Разумеется, это чисто теоретический вариант развития процесса происхождения человека.
Также интересным представляется то обстоятельство, что при росте общей численности популяции влияние количества «перекрестных браков» на численность прямых предков уменьшается. При количестве особей в сотни и тысячи, значение «перекрестных браков» в качестве фактора ограничивающего рост генетической базы теряет смысл. Всегда можно будет найти много пар особей, проживающих в коммутативной зоне, не имеющих прямой генетической связи в нескольких поколениях и даже в десятках поколений. Видимо существует минимальная численность группы особей, при которой такое влияние нужно учитывать. Исходя из изложенных выше соображений, полагаем, что эта численность составляет менее 256 особей.
Кроме того представляется, что верхний предел численности предков также существует. Невозможно представить себе контакты в виде «браков» между очень удаленными территориально группами одного вида в историческое время. Неизбежно происходит их обособление с началом формирования отличительных признаков, например, выделение отдельных рас внутри вида (черная, белая, желтая, красная расы человечества). Это похоже на формирование отдельных ветвей у дерева. В настоящее время в связи с глобализацией происходит стремительное объединение, перемешивание генетического материала человечества, но в прошлом изоляция отдельных групп ограничивала число возможных предков по нашей оценке до значений порядка нескольких тысяч особей в одном поколении (4-5 тысяч в одной «ветке»).
Во времена «переселения народов», войн, количество коммутаций между отдельными группами возрастает, в том числе количество «браков» с расширением генезисной базы. Генетические линии перемешиваются, «ветки» перекрещиваются. В периоды успокоения, освоения территорий и закрепления на достигнутых рубежах происходит обособление отдельных групп, вплоть до изоляции, что безусловно влияет на уменьшение генезисной базы.
Очевидно, что реальный процесс передачи генетической информации может иметь периоды экспоненциального роста численности участников этого процесса, периоды стабилизации численности, периоды умеренного роста, периоды сокращения за счет «перекрестных браков».
Если рассматривать процесс генезиса в прямом «прогрессивном» направлении (восходящем от прошлого к будущему), то следует учесть неуклонный рост общей численности населения Земли в течение последних тысячелетий, надежно подтвержденный данными науки. Увеличение общей численности населения можно рассматривать как расширение возможной генетической базы человечества и отдельного человека. Обратный «регрессивный» процесс как мы показали, тоже идет в общем направлении расширения генетической базы. Следовательно, неизбежно их пересечение, когда число предков сравняется с числом проживающих в ареале обитания, возможно со стабилизацией численности прямых предков в течение длительного времени. Все проживающие в ареале будут моими прямыми предками!
Графически это можно представить в виде «дерева жизни». Сначала снизу идет ствол из небольшого числа членов популяции, затем их число увеличивается, ствол распадается на отдельные ветки, затем некоторые ветки стабилизируется в росте, другие продолжают расширение или сужение, переплетаются между собой, наконец, формируется зонтичная вершина, сводящаяся к одному конкретному человеку.
Заключение.
У автора сложилось ощущение, что современная биология и генетическая биология в частности, находится в тупике. Эта наука опирается на фундаментальные законы, сформулированные еще в XIX-м веке Грегором Менделем. За прошедшие сто лет она научилась применять самые современные методы научного анализа, работать со сложными компьютерными программами, сложнейшими электронными приборами. Это позволило добиться определенных успехов, в частности в расшифровке генома человека, механизмов информационно-энергетического обмена в клетках. Сила биологии в ее системном подходе к самым разноплановым явлениям природы. Однако ее сила является и ее слабостью. Уязвимость биологии заключается в ее феноменологическом (описательном) характере. Она отвечает на вопрос «как?» но не отвечает на вопрос «почему?». Для этого необходимо от описания «явлений» переходить к описанию «объектов». На мой взгляд, без синтеза наук, применения новых методов и подходов из других наук – физики, математики, дальнейшее движение не возможно.
Представляется перспективным дальнейший ход исследования в области генезиса человека производить, используя методологию и формализм теории автоматического управления (ТАУ), теории информации (ТИ) и логических функций.
Процесс передачи генетической информации можно определить как дискретный сигнал со случайными шумами. Приемниками и передатчиками генетической информации являются организмы отдельных особей, даже еще конкретнее – хромосомы в половых клетках. Сразу отметим, что структура организмов особей нас не интересует.
Пользуясь терминологией ТАУ, можно рассматривать эти особи как «черный ящик», имеющий некую передаточную функцию для информационных сигналов. Передаточная функция включает внутренний набор собственных сигналов – последовательности генов ДНК особи. Эти сигналы можно рассматривать как генерацию внутреннего сигнала. На вход системы поступает закодированный сигнал в виде последовательности генов в ДНК другой особи, а на выходе получается суперпозиция входного сигнала и передаточной функции, при этом выходной сигнал формируется как вероятностная величина. Суперпозиция означает вероятность получения выходного сигнала из входного и сигнала внутреннего генератора сложением их по амплитудам, фазам и частотам (АФЧ - характеристика).
Например, известное выражение для пропускной способности канала связи передачи информации (формула Шеннона - Хартли):
C = W*log (1 + P/N), где:
С - пропускная способность канала связи (скорость передачи данных),
W – полоса частот сигнала (ширина спектра)
P – мощность сигнала
N – мощность шума
Важно отметить, что передаточная функция может иметь вид положительной либо отрицательной связи.
Сигнал, поступивший на вход, либо усиливается, либо ослабляется. Накопление мутаций – положительная связь, подавление мутаций – отрицательная связь. Этим понятиям можно сопоставить привычные для биолога понятия доминантных и рецессивных генов.
Какие известные биолого–генетические показатели можно сопоставить АФЧХ? На наш взгляд это следует сделать для корректности информационного представления рассматриваемого процесса передачи генов от поколения к поколению. Тогда могут быть применены методология и формализм теории информации. Определение этого соответствия понятий ТАУ и ТИ с традиционными биологическими понятиями является задачей следующих исследований.
Володин Владимир Иванович