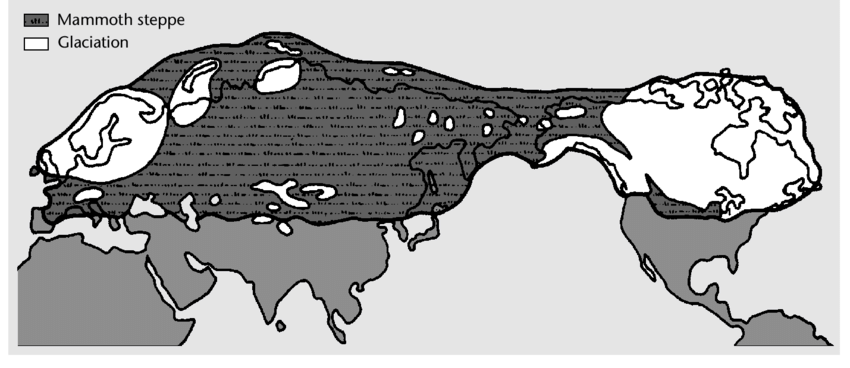
Происхождение индоевропейцев связано с приледниковой мамонтовой степью, которая в эпоху позднего палеолита простиралась от атлантического побережья Западной Европы до Северной Америки, соединённой тогда с Евразией сушей Берингии. Индоевропейский язык в своей ранней форме образовался ок. 5000 г. до н.э. на юге Русской равнины в среде носителей преимущественно мужских гаплогрупп R1a и R1b. Самым ранним известным представителем предковой для них обеих гаплогруппы R* (R-M207*) является мальчик, живший 24 тысячи лет назад на палеолитической стоянке Мальтá в Иркутской области к западу от Байкала. Его женская гаплогруппа U* была больше всего распространена у европейских охотников-собирателей позднего палеолита и мезолита. По аутосомным генам мальчик из Мальты был наиболее сходен с современными популяциями Южной Азии (37%), Европы (34%) и американскими индейцами (26%), причём из европейцев он был ближе всего к популяциям Северо-восточной Европы (которые унаследовали больше всего генов от европейских охотников-собирателей позднего палеолита и мезолита).

Близости к современным восточным азиатам и сибирским народностям геном мальчика из Мальты не имеет. При этом современные американские индейцы унаследовали от популяции, к которой он принадлежал, от 14 до 38% своих генов.[1] Отметим, что у американских индейцев преобладает ряд линий мужской гаплогруппы Q, являющейся «братской» для гаплогруппы R мальтинского мальчика. По всей видимости, носители R и Q какое-то время составляли единую популяцию. Потом носители Q отделились, где-то в Восточной Сибири смешались с пришедшими с юга восточными азиатами, после чего через Берингию переселились в Америку и дали начало американским индейцам.
Таким образом, выясняется, что уже в глубокой древности протоевропейское население, предковое для современных европейцев, особенно северо-восточных, было распространено далеко на восток, по меньшей мере до озера Байкал. Позднее его сменили пришедшие откуда-то с юга, вероятно, с территории Китая, восточные азиаты, ставшие предками современных сибирских народностей. Причём произошло это довольно недавно, что показывают исследования останков людей, живших 17 тысяч лет назад на стоянке Афонтова Гора на левом берегу Енисея в пределах современного города Красноярска. Сведения об отцовской гаплогруппе мужчины с Афонтовой Горы 2 противоречивы[2], а женщина с Афонтовой Горы 3 имела материнскую гаплогруппу R1b (не путать с имеющей такое же обозначение мужской гаплогруппой!). По аутосомным генам они оба близки к мальчику из Мальты и образовывают с ним общий кластер.[3] Входившие в этот кластер люди получили у генетиков наименование «древние северные евразийцы» (англ. Ancient North Eurasians, ANE).
Женщина с Афонтовой Горы 3 оказалась самой ранней известной носительницей гена светлых волос, который позднее был распространён индоевропейцами в Европе: «Производный аллель KITLG SNP rs12821256, который связан со светлым цветом волос у европейцев (и, вероятно, является его причиной), присутствует у охотников-собирателей из Самары и Муталы и с Украины (I0124, I0014 и I1763), а также у нескольких более поздних лиц, имеющих степных предков. Поскольку этот аллель обнаружен в популяциях, происходящих от восточных, а не от западных охотников-собирателей, его источником, вероятно, является популяция древних северных евразийцев (ANE). Подтверждением этого служит то, что самым ранним известным человеком с таким производным аллелем является [сибирская] северная евразийка со стоянки Афонтова Гора 3, которая напрямую датируется примерно 16 130 – 15 749 гг. до н.э. калибр. (14 710 ± 60 лет назад)».[4]
Языковым отражением генетической общности «древних северных евразийцев» может быть выделяемая рядом лингвистов «митийская» (англ. Mitian) группа языковых семей, именуемая так по форме местоимений первого и второго лица (*mi, *ti – ср. рус. мы, ты), которые для неё реконструируются. В митийскую группу включаются индоевропейские, уральские, тюркские, монгольские, тунгусские, юкагирские, нивхский, чукотско-камчатские и эскимосско-алеутские языки, и её распространение, таким образом, примерно совпадает с приледниковой мамонтовой степью позднего каменного века. Ближайшей к индоевропейской языковой семьёй является уральская, объединяемая иногда с ней в единую индоуральскую семью. Языковые предки уральцев были, по всей вероятности, непосредственными восточными соседями языковых предков индоевропейцев. Их разделение должно было произойти в конце последней ледниковой эпохи, когда образовавшееся от таяния ледника Хвалынское море (14-9 тыс. лет до н.э.), достигавшее на северо-востоке широты нынешнего Оренбурга, создало между ними естественную преграду. Индоуральцы, оказавшиеся западнее неё, на юге Русской равнины, стали индоевропейцами, оказавшиеся восточнее, на юге Западной Сибири, – финноуграми.
Из этого следует, что бывшие предками индоевропейцев носители гаплогруппы R* или уже её отдельных ветвей R1a и R1b должны были поселиться на Русской равнине ещё до окончания последней ледниковой эпохи. Однако они не были её первыми обитателями. Об их предшественниках нам известно благодаря исследованиям останков палеолитических людей со стоянок Костёнки (Воронежская область) и Сунгирь (Владимирская область). Мужчина из Костёнок 14 (Маркина Гора), живший примерно 36 тысяч лет назад, имел мужскую гаплогруппу C1b и женскую гаплогруппу U2. Мальчик из Костёнок 12, живший примерно на пять тысячелетий позже, имел такую же женскую гаплогруппу U2 и мужскую гаплогруппу CT.[5]
На стоянке Сунгирь в самом богатом на сегодняшний день в мире палеолитическом захоронении были обнаружены останки четырёх человек, живших 34 тысячи лет назад. Недавно выяснилось, что дети из двойного захоронения, раньше считавшиеся братом и сестрой, были оба мальчиками, причём не близкими родственниками. Старший мальчик приходился близким родственником похороненному неподалёку могучему мужчине, который при жизни занимался колкой камня. Они оба в течение жизни обильно питались мясом, а младший мальчик мяса вообще не ел, питаясь только беспозвоночными. Рядом с ним нашли человеческую кость, служившую ёмкостью для охры. Эта кость принадлежала близкому предку (предположительно прадеду) младшего мальчика, пища которого при жизни тоже состояла исключительно из беспозвоночных. По-видимому, они оба происходили из семьи шаманов. Старший мужчина был убит выстрелом из лука, а младший мальчик – ударом в живот. В их племя, судя по кругу семейных связей, входило 250-450 человек. Все четверо мужчин из Сунгири были носителями мужской гаплогруппы С1а2, при этом трое из них имели ту же женскую гаплогруппу U2, что и костёнковцы, а один – U8c.[6]
Таким образом, все костёнковцы и сунгирьцы были носителями ветвей женской гаплогруппы U, предковая линия которой была обнаружена у мальчика из Мальты. Что касается их мужских линий, то они в эту раннюю эпоху были, по всей видимости, широко распространены в Европе. Гаплогруппа CT мальчика из Костёнок 12 обнаружена также у его примерных современников – людей из Чокловины в Румынии (33 тысячи лет назад) и Вестонице в Чехии (31 тысяча лет назад). Гаплогруппа сунгирьцев C1a найдена также у их примерных современников – людей из Гойе в Бельгии (35 тысяч лет назад, C1a) и Павлова в Чехии (31 тысяча лет назад, С1а2), а, кроме того, у гораздо более позднего охотника-собирателя из Ла-Браньи в Испании (1-я пол. 8 тыс. до н.э., С1а2).[7]
Точное время и обстоятельства замещения на Русской равнине носителей гаплогрупп CT и C1 пришедшими с востока носителями гаплогруппы R на данный момент остаются неизвестными. Самым ранним свидетельством западной экспансии последних является живший ок. 14 тысяч лет назад охотник-собиратель из Виллабруны на северо-востоке Италии, имевший мужскую гаплогруппу R1b-L754 (R1b1a).[8] Та же линия была обнаружена у семи охотников-собирателей, живших между 9500-5900 гг. до н.э. в районе карпатских Железных ворот на границе Сербии и Румынии (ещё у десяти живших там же охотников-собирателей была обнаружена гаплогруппа I2a),[9] и у неолитического земледельца, жившего ок. 7 тысяч лет назад в Испании[10]. Однако эта представляющая раннюю западную экспансию R1b линия, по всей видимости, впоследствии вымерла и не оставила потомков, а все современные носители R1b в Западной Европе принадлежат к линии, которая была принесена индоевропейцами с юга Русской равнины в III тыс. до н.э.
Древнейшим известным в настоящее время представителем гаплогруппы R1a является восточноевропейский охотник-собиратель, живший ок. 8825-8561 гг. до н.э., останки которого были обнаружены в могильнике Васильевка 3, находившемся на левобережье Днепра близ порогов (нынешняя Днепропетровская область). Другой мужчина из того же могильника (живший ок. 8280-7967 гг. до н.э.) был носителем гаплогруппы I2a1, а мужчина из соседнего могильника Васильевка 2 (живший ок. 7446-7058 гг. до н.э.) – гаплогруппы R1b1a. У всех троих была обнаружена материнская гаплогруппа U5b2. Люди, захороненные в Волненском могильнике также в районе днепровских порогов (6500-4000 гг. до н.э.), были носителями гаплогрупп IJ, I, I2, I2a2, I2a2a и I2a2a1b1, ещё один человек с могильника Вовниги в нынешней Днепропетровской области (5473-5326 гг. до н.э.) – гаплогруппы I2a2a1b1b. Судя по отцовским линиям, они были пришельцами с запада, что также подтверждает примесь западноевропейских охотников-собирателей в их аутосомных генах. С материнской стороны у них были представлены разные линии гаплогрупп U2, U4 и U5. Люди из неолитического могильника у села Дереивка нынешней Кировоградской области на правом берегу Днепра (5500-4800 гг. до н.э.) были носителями мужских гаплогрупп R (2), R1b1 (1), R1b1a (5), R1a (1) и I2a2a1b (2) и женских гаплогрупп U4 и U5.[11]
В VI тысячелетии до н.э., на заре собственно индоевропейской истории, гаплогруппы R1a и R1b обнаруживаются у мезолитических охотников-собирателей в разных местах Русской равнины. R1a1a1 была найдена у человека, жившего ок. 5500-5000 гг. до н.э. (предлагаются и более ранние датировки) на Южном Оленьем острове Онежского озера, а R1b1a1a – у его примерного современника, жившего на реке Сок в нынешней Самарской области.[12] Носителем R1a был человек культуры Ямочно-гребенчатой керамики, живший в середине IV тыс. до н.э. в Кудрукюле на северо-востоке Эстонии.[13] Хотя он жил позднее интересующей нас эпохи, и его линия (R1a5-YP1272), как и линия человека из Карелии, является побочной для собственно индоевропейской, он свидетельствует о присутствии R1a у охотников-собирателей севера Русской равнины уже в довольно раннее время. R1b-L754 (R1b1a) также была обнаружена у шести представителей Нарвской культуры из могильника Звейниеки на севере Латвии, живших между 7465 и 4852 гг. до н.э. (ещё шесть человек из того же могильника оказались носителями I2a и один – Q1a2).[14]
Именно в среде таких перешедших к скотоводству мезолитических охотников-собирателей на рубеже VI-V тыс. до н.э. и зарождается на юге Русской равнины праиндоевропейский язык.
[1] Maanasa Raghavan et al. Upper Palaeolithic Siberian Genome Reveals Dual Ancestry of Native Americans // https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4105016/.
[2] Сначала было объявлено о R1a, потом – о Q1a1, позднее и эта информация была удалена из публикации.
[3] Qiaomei Fu et al. The Genetic History of Ice Age Europe // https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4943878/.
[4] “The derived allele of the KITLG SNP rs12821256 that is associated with – and likely causal for – blond hair in Europeans is present in one hunter-gatherer from each of Samara, Motala and Ukraine (I0124, I0014 and I1763), as well as several later individuals with Steppe ancestry. Since the allele is found in populations with EHG but not WHG ancestry, it suggests that its origin is in the Ancient North Eurasian (ANE) population. Consistent with this, we observe that earliest known individual with the derived allele is the [Siberian] ANE individual Afontova Gora 3 which is directly dated to 16130-15749 cal BCE (14710±60 BP)”: Iain Mathieson et al. The Genomic History Of Southeastern Europe // https://www.biorxiv.org/content/early/2017/09/19/135616.
[5] Qiaomei Fu et al. The Genetic History of Ice Age Europe // https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4943878/; Martin Sikora et al. Ancient Genomes Show Social and Reproductive Behavior of Early Upper Paleolithic Foragers // http://science.sciencemag.org/content/358/6363/659.
[6] Qiaomei Fu et al. The Genetic History of Ice Age Europe // https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4943878/; Martin Sikora et al. Ancient Genomes Show Social and Reproductive Behavior of Early Upper Paleolithic Foragers // http://science.sciencemag.org/content/358/6363/659.
[7] Maanasa Raghavan et al. Upper Palaeolithic Siberian Genome Reveals Dual Ancestry of Native Americans // https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4105016/.
[8] Qiaomei Fu et al. The Genetic History of Ice Age Europe // https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4943878/.
[9] Gloria Gonzalez-Fortes et al. Paleogenomic Evidence for Multi-generational Mixing between Neolithic Farmers and Mesolithic Hunter-Gatherers in the Lower Danube Basin // https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5483232/; Vagheesh M. Narasimhan et al. The Genomic Formation of South and Central Asia // https://www.biorxiv.org/content/10.1101/292581v1.
[10] Wolfgang Haak et al. Massive Migration from the Steppe is a Source for Indo-European Languages in Europe // https://www.biorxiv.org/content/early/2015/02/10/013433.
[11] Iain Mathieson et al. The Genomic History Of Southeastern Europe // https://www.biorxiv.org/content/10.1101/135616v1.supplementary-material?versioned=true.
[12] Wolfgang Haak et al. Massive Migration from the Steppe is a Source for Indo-European Languages in Europe // https://www.biorxiv.org/content/early/2015/02/10/013433.
[13] Lehti Saag et al. Extensive Farming in Estonia Started through a Sex-biased Migration from the Steppe // https://www.biorxiv.org/content/biorxiv/early/2017/03/02/112714.full.pdf.
[14] Eppie R. Jones et al. The Neolithic Transition in the Baltic was not Driven by Admixture with Early European Farmers // https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5321670/; Vagheesh M. Narasimhan et al. The Genomic Formation of South and Central Asia // https://www.biorxiv.org/content/10.1101/292581v1.