ТАКСОНЫ
Сдерживающие отношения между коронованными птицами в соответствии с принятыми отношениями качественно не изменили результатов. Аналогичным образом, повторение анализа после исключения таксонов подстановочных знаков существенно улучшает поддержку по всему дереву; однако это улучшение разрешения едва ли повлияло на оценку даты расхождения. Даты, найденные здесь, особенно чувствительны к включению или исключению форм палеогена. За исключением Gallinuloides (∼55 Ma), Lithornis (∼55.8 Ma) и Waimanu (∼59 Ma) публикуют гораздо более ранние прогнозные даты для кроны Авеш (∼60 Ma), которые датируются границей K-Pg.

В этих анализах верхний меловой вегавис извлекается в виде стебля, а не в виде кроновой птицы. Поэтому три палеогеновых пещеры имеют решающее значение для получения среднемеловой даты расхождения: производная морфология этих окаменелостей наиболее согласуется с относительно длительной историей эволюции пещерных пещер, которая простирается вглубь мелового пояса. Тем не менее, одно из основных потенциальных осложнений плохо аккомодатируется общими моделями часов, используемыми в датировке расхождений: оценка неоднородности во времени.
Например, если бы морфологические (и молекулярные) скорости были повышены по всем птичьим линиям во время взрывной ранней палеогенной диверсификации, даты расхождений, оцененные морфологическими (и молекулярными) методами синхронизации, были бы смещены в сторону более старых значений. Большие морфологические (и молекулярные) различия между современными линиями птиц ошибочно объясняются длительным периодом "нормального темпа" расхождения, а не более коротким периодом более быстрой эволюции. Длина ветвей в скупости и недатированные байесовские анализы также показывают, что более молодые таксоны последовательно подвергались более эволюционным изменениям, в соответствии с ожиданиями часовых моделей и общими данными, датируемыми стратиграфически более молодыми таксонами, должны (в среднем) иметь более анагенетический характер. Эта связь между возрастом и анагенезом имеет большое значение, используют ли они филогенетически некорректные или скорректированные сравнения. Например, существует весьма значительная связь между стратиграфическим возрастом таксонов на терминалах и их общим анагенезом, что отражается в длине пути от корня к кончику, но эти сравнения усложняются филогенетической ненезависимостью.
Стандартные сравнительные методы неприменимы к этим данным, поскольку ни "стратиграфический возраст", ни "тотальный анагенез" не удовлетворяют эволюционным моделям в таких методах, например, эти кумулятивные признаки не развиваются броуновским движением. Филогенетически независимые сравнения возможны, если сравнивать длительность каждой ветви с предполагаемой величиной эволюции, что приводит к значительной ассоциации.
Вероятность и байесовские подходы к томорфологической филогенетике находятся в относительном зачаточном состоянии и остается множество методологических вопросов; например, целесообразность применения какой-либо одной вероятностной (или паримониальной) модели к признакам с весьма неоднородной эволюционной динамикой, коррекция смещения при установлении адекватно компенсирует недостаточность выборки аутапоморфий и то, является ли морфологическое развитие слишком эпизодическим для применения даже расслабленных моделей.
Настоящий анализ является первой попыткой оценить морфологические - палеонтологические данные с количественными методами филогенетического знакомства, аналогичными тем, которые используются в молекулярных исследованиях, и следует надеяться, что он послужит стимулом для дальнейшего эмпирического анализа, особенно среди палеобиологов, который поможет найти ответы на эти вопросы. Молекулярный и морфологический подходы с использованием моделей часов имеют взаимодополняющие преимущества: молекулярные исследования могут анализировать огромное количество признаков с помощью хорошо проверенных стохастических моделей, в то время как морфологические исследования могут включать в себя более широкие выборки по промежуточным таксонам (вымершие формы) и по времени (стратиграфический возраст).
Эти два подхода предполагают в целом последовательные (среднемеловые) временные рамки эволюции для диверсификации современных птиц и предполагают, что многие линии современных птиц существовали в верхнем меловом поясе и пересекали границу КПГ. Если это так, то общий низкий потенциал ископаемого окаменелости птиц сам по себе недостаточен для объяснения отсутствия современных птиц до самого верхнего мелового периода, но дополнительным фактором может быть особенно неравномерная запись ископаемых верхнего мелового периода на южных континентах, где они могли возникнуть, и это приводит к значительной ассоциации.
Вероятность и байесовские подходы к томорфологической филогенетике находятся в относительном зачаточном состоянии и остается множество методологических вопросов; например, целесообразность применения какой-либо одной вероятностной (или паримониальной) модели к признакам с весьма неоднородной эволюционной динамикой, коррекция смещения при установлении адекватно компенсирует недостаточность выборки аутапоморфий и то, является ли морфологическое развитие слишком эпизодическим для применения даже расслабленных моделей.
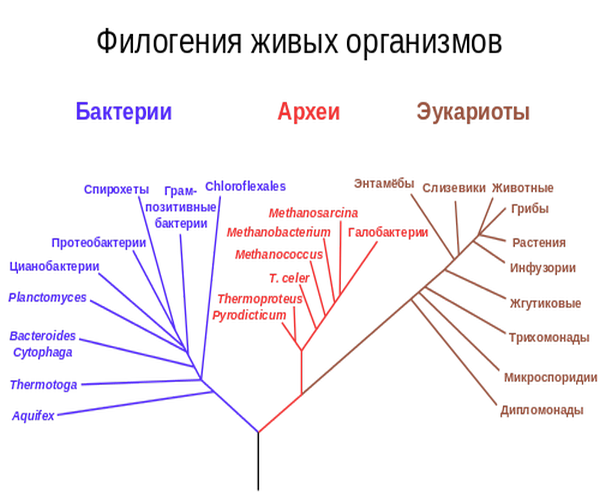
АНАЛИЗ
Настоящий анализ является первой попыткой оценить морфологические - палеонтологические данные с количественными методами филогенетического знакомства, аналогичными тем, которые используются в молекулярных исследованиях, и следует надеяться, что он послужит стимулом для дальнейшего эмпирического анализа, особенно среди палеобиологов, который поможет найти ответы на эти вопросы. Молекулярный и морфологический подходы с использованием моделей часов имеют взаимодополняющие преимущества: молекулярные исследования могут анализировать огромное количество признаков с помощью хорошо проверенных стохастических моделей, в то время как морфологические исследования могут включать в себя более широкие выборки по промежуточным таксонам (вымершие формы) и по времени (стратиграфический возраст).
Эти два подхода предполагают в целом последовательные (среднемеловые) временные рамки эволюции для диверсификации современных птиц и предполагают, что многие линии современных птиц существовали в верхнем меловом поясе и пересекали границу КПГ. Если это так, то общий низкий потенциал ископаемого окаменелости птиц сам по себе недостаточен для объяснения отсутствия современных птиц до самого верхнего мелового периода, но дополнительным фактором может быть особенно неравномерная запись ископаемых верхнего мелового слоя на южных континентах, где они могли возникнуть.