Всех приветствую на моем канале!!!
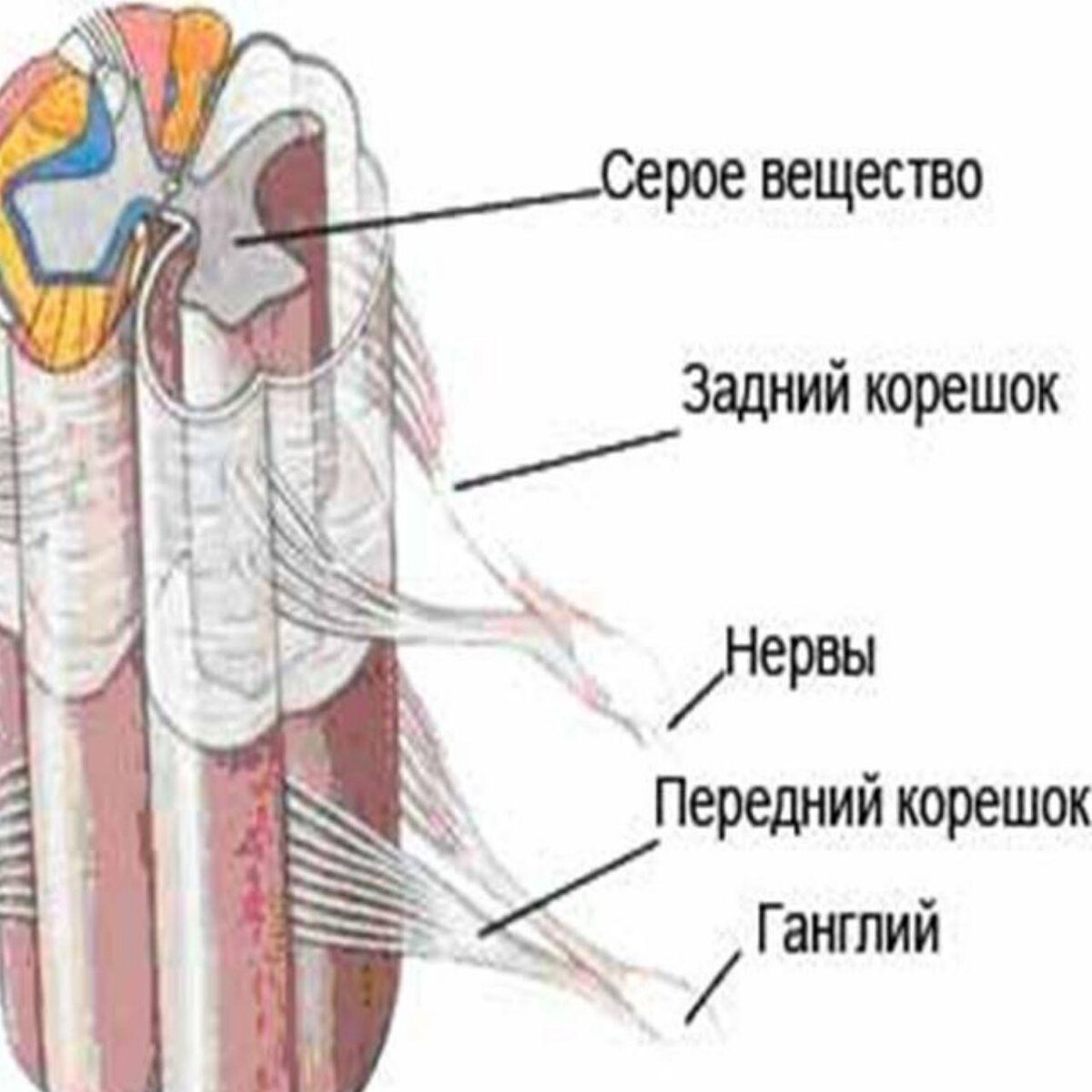
В условиях хронической боли повторяющиеся или интенсивные раздражители приводят к дезадаптивным пластическим изменениям вдоль пути боли , включая сенсибилизацию ноцицептивных цепей позвоночника, явление, известное как центральная сенсибилизация. Центральная сенсибилизация считается ключевым механизмом, лежащим в основе развития состояний постоянной гиперчувствительности. Изменения в некоторых клеточных процессах могут способствовать центральной сенсибилизации, включая усиление постсинаптического ответа спинномозговых нейронов на высвобождение нейротрансмиттера от первичных афферентов, снижение ингибирующего тонуса в результате снижения возбудимость спинномозговых ингибиторных интернейронов и неэффективная GABAergic и глицинергическая нейротрансмиссия , модуляция нисходящих путей.Дисбаланс возбуждающей и ингибирующей активности при центральной сенсибилизации приводит к повышенной возбудимости спинальной ноцицептивной схемы, что вызывает усиление периферического сигнала. Центральная сенсибилизация приводит к снижению болевого порога (аллодиния), увеличению воспринимаемой реакции на вредные раздражители (гипералгезия) и рекрутированию периферических сигналов из неповрежденных областей, что вызывает расширение рецептивного поля (вторичная гипералгезия).
Поступательный контроль нейрональной пластичности
Длительная модуляция внутренней возбудимости и синаптических функций зависит от экспрессии нового гена. Экспрессия гена может быть модулирована на различных этапах: транскрипции, мРНК перевода, мРНК и стабильности белка , и пост-трансляционной модификации белка. Трансляционный контроль позволяет модулировать клеточный протеом, регулируя эффективность, с помощью которой мРНК транслируется в белки. Он предоставляет нейронам механизм для быстрого и локального ответа на внутриклеточные стимулы и внеклеточные сигналы путем модификации их клеточного или синаптического протеома(совокупность белков организма,производимых клеткой,тканей в определенный момент времени).
Механизмы трансляционного контроля
Трансляция мРНК может быть разделена на три этапа: начало, удлинение и окончание. Инициирование является ограничивающим скорость шагом для перевода и поэтому жестко регулируется несколькими механизмами ( Sonenberg and Hinnebusch, 2009 ). На своем 5'-конце все транскрибируемые в ядре эукариотические мРНК содержат структуру, называемую 7-метилгуанозинтрифосфатом (m 7 Gppp), называемую «шапка». Эта структура облегчает рекрутирование рибосом в мРНК . 3'-конец мРНК содержит поли (А) хвост, который защищает мРНК от деградации и связывает поли (А) -связывающий белок (ПАБП). Механизмы, регулирующие инициацию переводаможно разделить на две основные категории: (1) регуляция рекрутирования рибосомы в колпачок на 5'-конце мРНК (посредством фосфорилирования факторов инициации трансляции, таких как 4E-BP, eIF4E и eIF2a), и (2) регуляция трансляции на 3'-конце мРНК путем контроля длины поли (А) хвоста (например, с помощью CPEB).
Для рекрутинга рибосомы требуется группы факторов eIF4 , который представляет собой комплекс из трех субъединиц, состоящий из eIF4A (РНК-геликазы),eIF4E, который специфически взаимодействует со структурным белком и eIF4G, большим каркасным белком, который связывается как с eIF4E, так и с eIF4A. eIF4G служит модульным каркасом, который собирает белковый механизм для направления рибосомы к мРНК .eIF4E обычно демонстрирует самый низкий уровень экспрессии всех эукариотических факторов инициации. Он играет центральную роль в распознавании головки, и из-за низкого уровня экспрессии он считается ограничивающим скорость шагом для перевода и главной целью для регулирования. Сборке eIF4F способствует механистическая мишень комплекса 1 рапамицина (mTORC1), который фосфорилирует и тем самым инактивирует трансляционные репрессоры , eIF4E-связывающие белки (4E-BP1)., 4E-BP2 и 4E-BP3). 4E-BP репрессируют образование комплекса eIF4F, конкурируя с eIF4G за общий сайт связывания на eIF4E. При фосфорилировании с помощью mTORC1 связывание 4E-BP с eIF4E снижается, что позволяет образовывать комплекс eIF4F и инициировать трансляцию. mTORC1 также фосфорилирует свои вторые главные нижестоящие эффекторы, S70 киназы p6 (S6K1 / 2), которые регулируют инициацию трансляции (через eIF4B), удлинение трансляции(через eEF2K) и биогенез рибосом (через рибосомный белок S6).
Активность eIF4E также регулируется с помощью фосфорилирования на серин 209 по MNK1 / 2 (митоген-активированной протеинкиназы (МАРК) , взаимодействующих протеинкиназ 1/2) ниже по потоку от ERK (внеклеточного сигнала регулируется киназы). Это событие фосфорилирования связано с повышением скорости инициации трансляции , хотя точный лежащий в основе молекулярный механизм остается неизвестным.
Второй основной механизм трансляционного контроля опосредуется фактором инициации трансляции, eIF2 (состоящим из трех субъединиц) , через фосфорилирование его α-субъединицы. Инициирование трансляции требует образования тройного комплекса, состоящего из инициатора (Met-tRNA i Met ) и связанного с GTP eIF2. В конце каждого раунда рекрутирования рибосом происходит рециркуляция неактивного eIF2ɑ, связанного с GDP, в активный eIF2, связанный с GTP, с помощью фактора обмена гуаниновых нуклеотидов (GEF), eIF2B . Фосфорилирование eIF2ɑ в серине 51 ингибирует активность eIF2B, уменьшая образование тройного комплекса и тем самым ингибируясинтез белка . Как это ни парадоксально, фосфорилирование eIF2ɑ стимулирует трансляцию мРНК, содержащих открытые открытые рамки считывания (uORF) в их 5'-UTR, таких как ATF4 и CHOP. eIF2ɑ фосфорилируется в ответ на различные клеточные стрессовые состояния посредством активации киназ eIF2ɑ (PERK, PKR, GCN2 и HRI) .Фосфорилирование eIF2ɑ в значительной степени участвует в регуляции общей трансляции, тогда как eIF4E-зависимый трансляционный контроль регулирует трансляцию отдельного подмножества мРНК, многие из которых участвуют в пролиферации, росте и синаптической пластичности .
Трансляция также регулируется с помощью 3'-опосредованных механизмов. Трансляция мРНК, содержащих элементы цитоплазматического полиаденилирования (CPE) на их 3'-UTR , регулируется белком, связывающим элементы цитоплазматического полиаденилирования (CPEB) . CPEB связывает CPE и стимулирует удлинение поли (A) хвоста, регулируя аппарат полиаденилирования, состоящий из поли (A) полимеразы Gld2, деаденилазы PARN и трансляционного фактора нейрогидина (Ngd). Удлинение поли (А) хвоста мРНК приводит к стабилизации мРНК и усилению связывания поли (А) -связывающего белка (ПАБП), что облегчает инициацию трансляции путем одновременного связывания как с хвостом поли (А), так и с eIF4G, в результате чего в циркуляции мРНК .Было показано, что этот механизм регулирует трансляцию мРНК CamkIIα и Nr2a .