Клеточные органеллы, на которых происходит синтез белка, построены из трех разных РНК и пятидесяти двух белков. Каким образом регулируется сборка рибосом в соответствии с нуждами клетки?
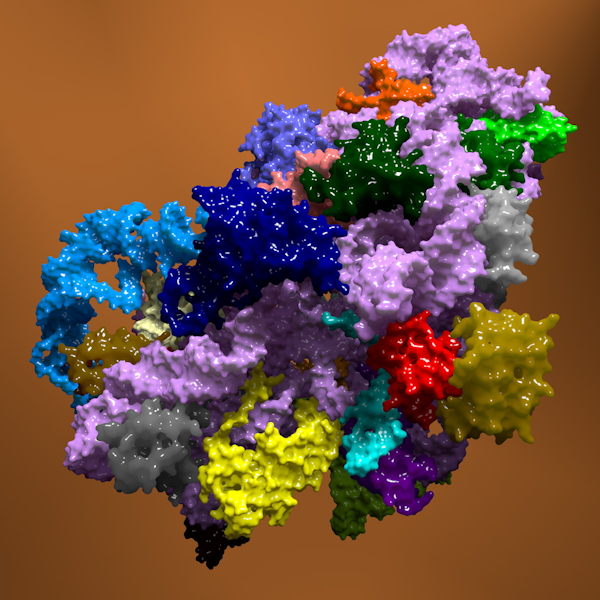
Рибосомы — это клеточные органеллы, осуществляющие синтез белков в клетке. На них происходит последовательное присоединение аминокислот к растущей полипептидной (белковой) цепи. Завершенная белковая цепь может сама сворачиваться в пространстве, становясь функционально активной молекулой, которая либо выполняет в клетке структурную функцию, либо служит ферментом, т.е. катализирует ту или иную биохимическую реакцию.
Так как все реакции обмена веществ протекают при участии ферментов, для того чтобы быстро расти, клетке необходимо ускорять биосинтез белков. Однако на рибосоме скорость присоединения новых аминокислот к растущей цепи белка ограничена.
К примеру, у бактерий “Escherichia coli” при 37°С на одной рибосоме может происходить присоединение не более 15 аминокислот в секунду. Если температура остается постоянной, а среда, в которой живет “E. coli”, обогащается питательными веществами, бактерии начинают делиться значительно быстрее.
Поскольку возможности отдельной рибосомы ограничены, клетке для ускорения синтеза белка необходимо иметь больше рибосом. Но на их биосинтез расходуется довольно много энергии, и, следовательно, иметь большой избыток рибосом для клетки тоже невыгодно. Значит, бактерии должны иметь достаточно тонкие механизмы контроля числа рибосом.
Рибосома устроена довольно сложно: в ее состав входят 52 различных белка и 3 разные РНК, так что и регуляция ее биосинтеза далеко не проста. Информация о структуре рибосомных белков и рибосомных РНК (р РНК) бактериальной клетки закодирована в ДНК ее единственной хромосомы.
При синтезе белков эта информация сначала «переписывается» (транскрибируется) в РНК, которая далее на рибосоме служит матрицей для сборки белков. р РНК же образуется непосредственно в результате транскрипции с хромосомной ДНК.
Для обеспечения нормальной скорости биосинтеза рибосом необходимо, чтобы скорость синтеза каждого компонента определялась по отношению к скоростям синтеза остальных 54 компонентов, точно так же, как для сборки автомобиля необходимо наличие всех деталей на складе и их своевременное поступление на конвейер.
Регуляция сборки рибосом может, очевидно, осуществляться по крайней мере на двух уровнях — при транскрипции всех 55 рибосомных компонентов и при трансляции м РНК 52 рибосомных белков.
Генетическое кодирование
Структура, функции и способ сборки рибосом настолько тесно связаны с экспрессией генетической информации, что необходимо сначала разобраться в генетическом кодировании, чтобы понять, как регулируется биогенез рибосом.
ДНК — первичный носитель генетической информации — представляет собой линейный полимер, в состав которого входят нуклеотиды четырех типов:
- аденин (А),
- цитозин (С),
- гуанин (G),
- тимин (Т).
Вследствие своей химической природы “А” обычно может соединяться водородными связями только с “Т”, а “С” — только с “G”.
Благодаря этой избирательности, или, как ее называют, комплементарности, две цепи ДНК соединяются в двунитевую спираль, такую, например, как единственная хромосома “E. coli”, несущая всю генетическую информацию о бактериальной клетке.
Хромосома “E. coli” представляет собой двуспиральную ДНК длиной около 4 млн. нуклеотидов, замкнутую в кольцо. В последовательности нуклеотидов закодирована информация о структуре всех белков, необходимых для бактерии, и некоторых ее РНК.
Участок ДНК, несущий информацию о каком-то одном продукте — белке или рибосомной РНК, называется геном. На гены 55 компонентов рибосомы приходится около 5% всей информации, заключенной в ДНК “E. coli”.
Последовательность нуклеотидов гена транскрибируется в последовательность нуклеотидов матричной РНК с помощью специальных ферментов — РНК-полимераз. РНК- полимераза связывается в определенном участке с ДНК и дальше движется по ней, синтезируя цепь РНК.
РНК состоит из тех же нуклеотидов, что и ДНК, за исключением того, что вместо тимина в нее входит урацил (U), который, так же как и тимин, комплементарен аденину. РНК-полимераза присоединяет к растущей цепи РНК нуклеотиды, комплементарные нуклеотидам в ДНК. Синтезированная РНК является, таким образом, «негативом» данного участка бактериальной хромосомы и, следовательно, несет ту же генетическую информацию, что закодирована в этом гене. По завершении транскрипции образовавшаяся молекула РНК отделяется от ДНК.
Генетические процессы
Какие же генетические процессы управляют синтезом рибосом в бактериальной клетке? Лучше всего изучены на сегодняшний день рибосомы “E. coli”, и в отношении их картина в целом уже довольно ясная.
Как и другие бактерии, “E. coli” относится к прокариотам; это означает, что у нее нет четко выраженного ядра. В прокариотических клетках транскрипция и трансляция тесно сопряжены.
Очень часто рибосомы начинают транслировать м РНК еще до того, как завершится ее транскрипция и она высвободится из комплекса с ДНК. По окончании транскрипции м РНК трансляция продолжается.
Однако, как правило, срок жизни м РНК невелик — всего несколько минут, после чего она быстро деградирует под действием определенных ферментов. За это время кодирующая последовательность успевает про транслироваться в среднем 10—20 раз.
Таким образом, на одной молекуле м РНК образуется несколько молекул белка. Количество синтезированных полипептидных цепей зависит от времени жизни м РНК и скорости трансляции. Многоклеточные организмы, в том числе и человек, относятся к эукариотам: их клетки имеют четко выраженное ядро, отделенное от окружающей цитоплазмы ядерной мембраной. В эукариотических клетках транскрипция происходит в ядре, а затем м РНК транспортируется в цитоплазму, где и осуществляется трансляция; сборка рибосом происходит в ядрышках — особых органеллах, расположенных в ядре клетки.
Рибосома в клетке
Первые измерения содержания рибосом в клетке основывались на определении количества РНК на разных стадиях роста и деления клеток.
В конце 50-х годов О.Маалу, Н.Кьельдаля и их коллеги в Государственном институте сывороток (Дания) и позже в Институте микробиологии Копенгагенского университета изучали клеточное содержимое бактерий “Salmonella typhimurium” — близких родственников “E. coli”.
Выращивая сальмонеллу на различных питательных средах, они обнаружили, что существует прямо пропорциональная зависимость между скоростью деления клеток и количеством РНК в них.
К этому времени уже было известно, что большая часть клеточной РНК приходится на долю рибосомной РНК, так что, измеряя содержание РНК, эти исследователи практически определяли концентрацию рибосом в бактериальной клетке.
Исследование рибосомных белков
Первым объектом исследования стали рибосомные белки. Примерно в то время, когда началось изучение проблемы регуляции, Е. Кальтшмидт и Х. Г. Виттман из Института молекулярной генетики им. Макса Планка в Западном Берлине разработали простой метод разделения практически всех рибосомных белков. Этот метод, а также другие, предложенные позже, позволили исследовать синтез индивидуальных рибосомных белков при различных скоростях роста клетки.
В 1974 г. П. Деннис в моей лаборатории в Висконсинском университете в Мадисоне применил новые методы разделения рибосомных белков для изучения их биосинтеза. Он показал, что скорости синтеза большинства белков сбалансированы между собой и что регуляция скоростей синтеза осуществляется скоординированно в ответ на изменение состава питательной среды, в которой росли клетки. Зная это, мы могли уже впрямую заняться проблемой регуляции.
С самого начала было очевидно, что изучение биосинтеза рибосом на молекулярном уровне малопродуктивно без выявления генов рибосомных белков и рибосомных РНК. Выделив эти гены, их можно было бы использовать для синтеза р РНК и рибосомных белков “in vitro” (т.е. «в пробирке») в бесклеточной системе.
Имея такую систему искусственного синтеза, вполне реально с помощью биохимических методов определить степень экспрессии индивидуальных генов, и понять механизм ее регуляции. Однако выделение генов 55 рибосомных компонентов оказалось довольно сложной задачей и заняло много времени. Методы рекомбинантных ДНК тогда еще не были разработаны, и приходилось опираться только на методы классической генетики.
Работа строилась в основном на манипулировании с бактериофагом “X” — вирусом, поражающим бактерии. При инфекции вирусная ДНК встраивается в хромосому клетки, после чего фаг может мирно сосуществовать с бактерией довольно долго.
Однако под влиянием некоторых факторов, например при повреждении клеточной ДНК в следствии облучения ультрафиолетом, фаг подавляет нормальную деятельность генетического аппарата клетки и начинается воспроизведение вирусных частиц.