Статодинамика взорвет объемы ваших мышц. Особенно эффективно на ноги! Ссылка на приложение Андроид --- а это ссылка на статью --->Ссылка >>
Определение ПАНО серьезная спортивно-научная проблема. По современным представлениям именно ПАНО решает кто на пьедестале, а кто бегает трусцой по утрам. Вот так примерно во всех работах выглядят графики pH крови и лактат как функция от мощностной нагрузки на спортсмена. В данном случае бег.
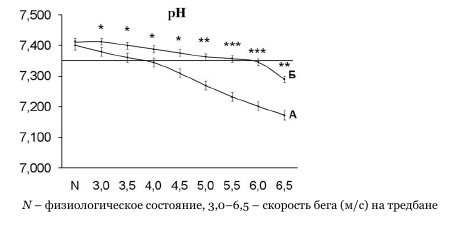
Эти данные интересны в заимосвязи с уровнем лактата в крови, который в свою очередь является основой формирования понятия ПАНО или АнП.
Мы видми на графиках точки излома. Их и принято считать точками входа на уровень выше ПАНО.
Результаты исследования
В состоянии покоя концентрация лактата в крови не отличалась у спортсменов первой и второй экспериментальных групп. Начиная со скорости 3,0 м/с и на всем протяжении эксперимента концентрация лактата в крови спортсменов увеличивалась у обеих экспериментальных групп. Однако у спортсменов высокой квалификации показатели были ниже, чем у начинающих спортсменов: на скоростях 3,0 м/с, 3,5 м/с, 4,0 м/с -приблизительно на 40%, на скорости 4,5 м/с - на 55%, на скоростях 5,0 м/с, 5,5 м/с, 6,0 м/с - примерно на 65% (рис. 1).
Концентрация ионов водорода увеличивается при увеличении скорости бега и повышении уровня лактата в крови. В состоянии покоя значения рН у спортсменов обеих экспериментальных групп были одинаковыми. Со скорости 3,0 м/с на всем протяжении эксперимента концентрация протонов у начинающих спортсменов была на 15-25% выше, чем у спортсменов высокой квалификации (рис. 1, А).
Парциальное давление углекислого газа рСО2 в физиологическом состоянии в крови спортсменов высокой квалификации было на 16% выше (рис. 2), чем у спортсменов первой группы, что приводило к повышению концентрации НСО3- и ВЕ у второй экспериментальной группы по сравнению с первой на 10% и 150% соответственно. Эти различия наблюдаются на всех этапах физической работы (рис. 2). Таким образом, увеличенные буферные резервы крови спортсменов второй группы обеспечивают высокую степень связывания протонов, образующихся при диссоциации лактата, и отодвигают время наступления порога анаэробного обмена. Кривые величин рСО2, НСО3- и ВЕ при увеличении интенсивности работы не содержат изломов и представляют собой линейные процессы.
Обсуждение результатов
При скорости бега 4,0 м/с у спортсменов первой экспериментальной группы концентрация лактата начинает резко возрастать с 4,1 ± 0,36 до 16 ± 1,2 ммоль/л (рис 1, А). Согласно гипотезе анаэробного порога, эта точка соответствует величине порога анаэробного обмена, что совпадает с данными, имеющимися в литературе [10, 11, 14, 20]. Однако, у спортсменов высокой квалификации концентрация лактата начинает резко возрастать с 4,6 ± 0,47 ммоль/л при скорости 5,0 м/с (рис 1, Б). Возрастание величины лактатного порога при адаптации к спортивным тренировкам достаточно хорошо описано [1, 2, 9-11] и предложены гипотетические молекулярные механизмы подобного сдвига [2, 4, 9]. Тем не менее Davis [10] показывает, что определение динамики изменения концентрации лактата не может служить методикой определения величины анаэробного порога, поскольку усиленный выброс лактата является лишь показателем уже совершившегося перехода метаболизма глюкозы на преимущественно гликолитический и не отражает причину начала повышенного образования лактата, а, соответственно, и истинную точку порога анаэробного обмена.
Практически все исследования в области метаболизма при физической работе оперируют предположением о том, что увеличение концентрации лактата связано с началом локальной мышечной гипоксии при определенной интенсивности работы [7, 8, 26]. Однако, многочисленные попытки установить связь между легочной вентиляцией, потреблением кислорода и лактатным порогом показали, что прямой связи нет [14, 15, 21], и, если процессы вентиляции и потребления кислорода изменяются линейно при линейно нарастающей интенсивности физической нагрузки, то и увеличение концентрации лактата должно происходить линейно, без точки излома.
Интересным является тот факт, что на кривой зависимости величины рН от интенсивности работы обнаруживается явный излом, точка, по времени предшествующая точке излома в концентрации лактата, - 4,0 м/с
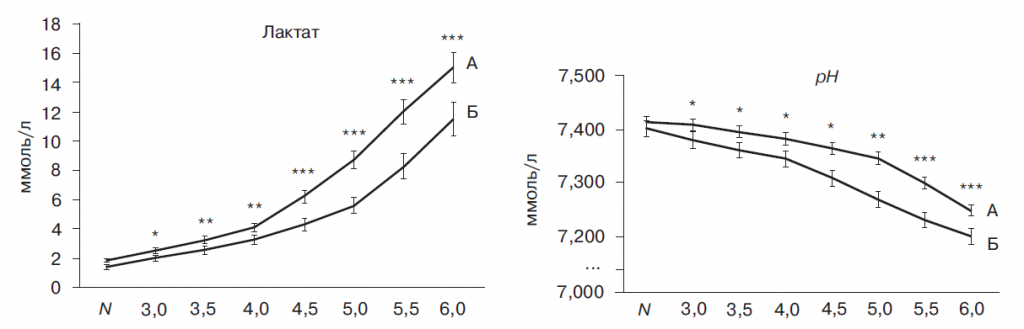
Рис. 1. Концентрация молочной кислоты (лактат, ммоль/л) и значение водородного показателя (рН) в крови спортсменов (n = 19-24) в норме и при физической работе
Рис. 2. Парциальное давление углекислого газа (рСО2, мм рт. ст.), концентрация гидрокарбонат-анионов (НСО/, ммоль/л) и концентрация избытка буферных оснований (ВЕ, ммоль/л) в крови спортсменов (n = 19-24) в норме и при физической работе
Пояснения к рис. 1 и 2:
А - первая экспериментальная группа, Б - вторая экспериментальная группа; * - P < 0,05, ** - P < 0,01, *** - P < 0,001.
По оси абсцисс отмечены: N - физиологическое состояние; 3,0-6,5 - скорость бега (м/с) на тредбане.
у первой экспериментальной группы и 5,0 м/с - у второй группы. Значение водородного показателя в этой точке соответствует нижней границе клинической нормы -рН = 7,35. Согласно литературным данным, пируватдегидрогеназный комплекс и ферменты цикла трикарбоновых кислот не активны при рН ниже физиологических значений [18]. Таким образом, можно предположить, что инактивация ферментов аэробного расщепления пировиноградной кислоты приводит к резкому увеличению концентрации лактата в мышечных клетках и крови, это в свою очередь приводит к появлению излома на кривой зависимости концентрации лактата от скорости бега (рис. 1). Ранее Wasserman [23] и Davis [9] указывали на ограниченную работоспособность систем, окисляющих пируват, как на возможную причину анаэробного порога. Однако, эта гипотеза не объясняла наличие излома на кривой зависимости концентрации лактата от интенсивности физической работы. К настоящему времени имеются сведения об увеличенной активности ферментов митохондрий у спортсменов высокой квалификации по сравнению с начинающими спортсменами [4, 9, 19].
Полученные результаты доказывают справедливость гипотезы лактатного порога и могут быть положены в основу разработки методики диагностики величины порога анаэробного обмена по водородному показателю.
Для демонстрации эффективности и универсальности предложенной нами методики была проведена работа по тестированию спортсменов специализации «плавание» в различных режимах работы. Было протестировано 2 группы пловцов (по 20 человек в каждой) квалификации «Мастер спорта» и «Мастер спорта международного класса». В ходе обследования определялись показатели рН и концентрации лактата крови.
Первая группа выполняла аэробно-анаэробный тест со ступенчато-повышающейся нагрузкой в виде 8 отрезков по 50 м, время отдыха между отрезками - 30 с, в это время производился отбор капиллярной крови из пальца. Перед тестируемыми спортсменами была поставлена задача плыть первый отрезок со скоростью 60% от максимальной, прибавляя скорость в каждом отрезке по 5%.
Результаты проведенного исследования показывают, что после 4 этапа со скоростью 75% от максимальной наблюдается достижение значения рН = 7,35, что сопровождается концентрацией лактата 6,01 ммоль/л. Данная концентрация лактата является характерной для точки ПАНО у высококвалифицированных спортсменов, и соотносится с результатами в тесте на тредбане (рис. 1).
Вторая группа спортсменов выполняла сходный тест в виде 6 отрезков по 100 м, время отдыха между отрезками - 60 с, в это время производился отбор капиллярной крови из пальца. Перед тестируемыми спортсменами была поставлена задача плыть первый отрезок со скоростью 60% от максимальной, прибавляя скорость в каждом отрезке по 5%.
Полученные результаты показывают сходные с предыдущим тестом величины рН и концентрации лактата в крови, несмотря на различающиеся длины отрезков и время работы в тесте и на каждом отрезке, что свидетельствует об универсальности методики, базирующейся не на измерении концентраций лактата, а на определении значения рН в тесте.
Заключение
Концентрация лактата в точке ПАНО не является постоянной величиной и зависит от степени тренированности спортсмена, а значение рН в точке ПАНО, как показывают результаты нашего исследования, всегда одинаковое - 7,35. Определение величины рН будет являться более универсальной методикой и не требовать объемной работы по построению графиков с определением точки излома в зависимости концентрации лактата от времени работы.
Литература/References
- Billat, V.L., Koralsztein, J.P., Morton, R.H. Differential modeling of anaerobic and aerobic metabolism in the 800-m and 1,500-m run J. Appl. Physiol. 107 (2): 478-487. 2009.
- Billat, V.L., Sirvent, P., Py, G, Koralsztein, J.P., and Mercier, J. The concept of maximal lactate steady state: a bridge between biochemistry, physiology and sport science. Sports Med. 33: 407-426. 2005.
- Brooks, G.A. Response to Davis' manuscript. Med. And Science in Sports and Exercise. 17 (1): 19-21. 1985.
- Brooks, G.A., Dubouchaud, H, Brown, M, Sicurello, J.P., Butz, C.E. Role of mitochondrial lactate dehydrogenase and lactate oxidation in the intracellular lactate shuttle. Pro. natl. Acad. Sci. USA. 96: 1129-1134. 1999.
- Brooks, G.A., Fahey, T.D. Exercise Physiology: Human Bioenergetics and its Applications. John Wiley and Sons, New York. 1984.
- Connett, RJ., Gaueski, T.E.J., Honig, C.R. Lactate accumulation in fully aerobic, working dog gracilis muscle. Am. J. Physiol. 246 (8): 120-128. 1984.
- Corbett, J., Barwood, M.J., Parkhouse, K. Effect of task familiarisation on distribution of energy during a 2000 m cycling time trial. Br. J. Sports Med. 43 (10): 770-774. 2009.
- Davis, H.A. and Gass, G.C. The anaerobic threshold as determined before and during lactic acidosis. Eur. J. Appl. Physiol. 47: 141-149, 1981.
- Davis, J.A. Anaerobic threshold. Med. and Science in Sports and Exercise. 17 (1): 1-18. 1985.
- Davis, J.A. Validation and determination of the anaerobic threshold. J. Appl. Physiol. 57 (1): 611. 1984.
- Davis, J.A. Does the gas exchange anaerobic threshold occur at a fixed lactate concentration of 2 or 4 mM? Int. J. Sports Med. 4 (4): 89-93. 1983.
- Douglas, C.G. Coordination of the respiration and circulation with variation in bodily activity. Lancet. 312 (1): 213-218. 1927.
- Gaw, A.A., Cowan, R.A., Stewar,t M.J., Sheperd, J. Clinical Biochemistry. Churchill Livingstone, Edinburg. 1999.
- Geir, S., Robstad, B., Skjensberg, O.H., Borchsenius, F. Respiratory gas exchange indices for estimating the threshold. Journal of Sports Science and Medicine. 4: 29-36. 2005.
- Green, H.J., Hughson, R.L., Orr, G.W., Ranney, D.A. Anaerobic threshold, blood lactate, and muscle metabolites in progressive exercise. J. Appl. Physiol. 54 (12): 1032-1038. 1983.
- Hetenyi, G., Perez, G., Vranic, M. Turnover and precursor product relationships of nonlipid metabolites. Physi- ol. Rev. 63: 606-667. 1983.
- Holtmann, W.F. Zur frange der dauerleistungsfahig-keit. Fortschr. Med. 7 (4): 443-453. 1961.
- Inui, H., Miyatake, K., Nakano, Y. and Kitaoka, S. Occurrence of oxygen-sensitive, NADP+-dependent pyruvate dehydrogenase in mitochondria. J. Biochem. 96 (1): 931-934. 1984.
- Perrey, S., Burnley, M., Millet, G.P., Borrani, F., Jones, A.M., Poole, D.C., Copp, S.W., Hirai, D.M., Gimenez, P., Bus-so, T. Comments on point: counterpoint: the kinetics of oxygen uptake during muscular exercise do/do not manifest time-delayed phase. Modeling concerns. J. Appl. Physiol. 107 (5): 1669-1670. 2009.
- Ramsbottom, R., Kinch, R.F., Morris, M.G., Dennis, A.M. Practical application of fundamental concepts in exercise physiology. Advan. Physiol. Educ. 31 (4): 347-351. 2007.
- Rees, B.B., Boily, P., Williamson, L.A. Exercise- and hypoxia-induced anaerobic metabolism and recovery: a student laboratory exercise using teleost fish. Advan. Physiol. Educ. 33 (1): 72-77. 2009.
- Robergs, R.A., Ghiasvand, F., Parker, D. Biochemsitry of exercise-induced metabolic acidosis. American Journal of Physiology: Regulatory, Integrative and Comparative Physiology. 287: 502-516. 2004.
- Wasserman, K.Y., Mcllroy, M.B. Detecting the threshold of anaerobic metabolism in cardiac patient during exercise. Am. J. Cardiol. 14 (3): 844-852. 1964.
- Westerblad, H., Allen, D., Jannergren, J. Muscle Fatigue: Lactic Acid or Inorganic Phosphate the Major Cause? News Physiol. Sci. 17: 17-21. 2002.