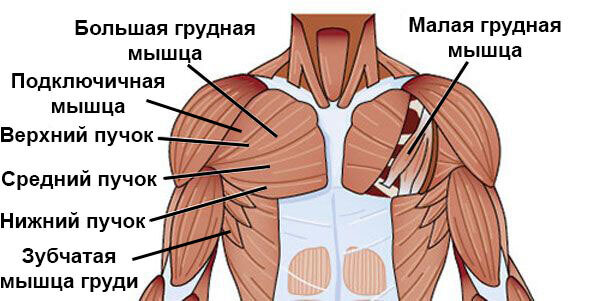
Одной из основной функции наружной мышечной системы
(соматической) – это передвижение тела человека в пространстве
(ходьба или бег) и осуществляется, как мы знаем, при помощи
нижних свободных конечностей ног (лат. mémbrum inférius
liberum), снабжённых самыми крупными мышцами (скелетными
мышцами) ног: мышц таза, мышц бедра и мышц голени (вместе с
мышцами пальцев ног).
Эти мышцы сокращаются усилием воли
самого человека под действием импульсов, поступающих к ним
по нервным волокнам из центральной нервной системы, и
обеспечивают возможность выполнения произвольных
движений, совершают мощные и быстрые сокращения, но
быстро утомляются. Скелетные мышцы (у человека их более 600)
прикреплены к костям или друг к другу крепкими, гибкими
сухожилиями, имеющими высокую прочность и низкую
растяжимость, образованны из соединительной ткани и являются
концевой структурой поперечно полосатых мышц, с помощью
которых через сухожилия они прикрепляются к костям скелета.
Скелетные мышцы сокращаясь воздействуют на кости или кожу,
к которым они прикрепляются, при этом один из пунктов
прикрепления остаётся неподвижным — так называемая точка
фиксации (лат. púnctum fíxum), которая в большинстве случаев
рассматривается в качестве начального участка мышцы, а
движующуюся точку прикрепления мышцы называют подвижной
точкой (лат. púnctum móbile). В каждом движении участвуют
мышцы как совершающие его, так и противодействующие ему
(агонисты и антагонисты соответственно), что придаёт движению
точность и плавность.
Самые сильные мышцы — икроножные и жевательные, а самая
длинная мышца (портняжная) начинается от передней верхней
ости крыла подвздошной кости (переднего верхнего отдел
тазовой кости), спиралевидно перекидывается спереди через
бедро и прикрепляется сухожилием к бугристости
большеберцовой кости (верхний отдел голени).
Поперечнополосатые мышечные ткани имеют две основные
разновидности поперечнополосатых (исчерченных) тканей —
скелетная мышечная ткань и сердечная мышечная ткань.
Функциональной единицей скелетной мышцы является моторная
единица (МЕ), включающая в себя группу мышечных волокон
(миоцитов) и иннервирующий их мотонейрон; число мышечных
волокон, входящих в состав одной МЕ, варьирует в разных
мышцах, например, там, где требуется тонкий контроль
движений (в пальцах или в мышцах глаза), МЕ небольшие, они
содержат не более 30 волокон. А в икроножной мышце, где
тонкий контроль не нужен, в МЕ насчитывается более 1000
мышечных волокон.
Моторные единицы одной мышцы могут быть разными: в
зависимости от скорости сокращения моторные единицы
разделяют на медленные/slow (S-МЕ) и быстрые/fast (F-МЕ), а
быстрые (F-МЕ) в свою очередь делят по устойчивости к
утомлению на быстроутомляемые/fast-fatigable (FF-МЕ) и
устойчивые к утомлению/fast-fatigue-resistant (FR-МЕ) и
соответствующим образом подразделяют иннервирующие
данные МЕ мотонейроны: S-мотонейроны(S-МН), FF-
мотонейроны (F-МН) и FR -мотонейроны(FR-МН).
Мотонейро́н (от лат. motor — приводящий в движение нейрон;
двигательный нейро́н) представляет из себя крупную нервную
клетку, расположенную в передних рогах спинного мозга, и,
обеспечивающую моторную координацию и поддержание
мышечного тонуса мышц человека, функционально различаются
на альфа-мотонейроны, иннервирующие волокна скелетной
мускулатуры (экстрафузальные волокна) и обеспечивающие
мышечное сокращение и гамма-мотонейроны, иннервирующие
рецепторы растяжения (интрафузальные волокна); мотонейроны
называются по той мышце, которую они иннервируют
(четырёхглавые, икроножные, полусухожильные и т. п.).
Скелетная мышечная ткань состоит из волокон миосимпласта и
миосателлитоцитов, покрытых общей базальной мембраной
(длина всего волокна может измеряться сантиметрами при
толщине всего 50—100 мкм, а комплекс, состоящий из
плазмолеммы миосимпласта и базальной мембраны, называют
сарколеммой.
Миосимпласт имеет множество продолговатых ядер,
расположенных непосредственно под сарколеммой (саркомер —
это структурная единица миофибриллы) и их количество в одном
миосимпласте может достигать нескольких десятков тысяч; у
полюсов ядер располагаются органеллы общего значения —
аппарат Гольджи и небольшие фрагменты гранулярной
эндоплазматической сети запасающие ионы кальция (Ca2+)
(саркоплазматический ретикулюм, митохондрии и др.);
миофибриллы заполняют основную часть миосимпласта и
расположены продольно; каждая миофибрилла имеет
поперечные темные и светлые диски, имеющие неодинаковое
лучепреломление (анизотропные A-диски и изотропные I-диски)
и окружена продольно расположенными и анастомозирующими
между собой петлями агранулярной эндоплазматической сети
(саркоплазматической сети или саркоплазматического
ретикулума); соседние саркомеры имеют общую пограничную
структуру — Z-линию (или телофрагму), построеную в виде сети
из белковых фибриллярных молекул связаных концами с
тонкими актиновыми филаментами, среди которых
существенную роль играет альфа-актинин, объединёный с Z-
линией и нитями миозина, фибриллярными нерастяжимыми
молекулами небулина, и фиксированы концами по отношению к
Z-линиям растяжимыми гигантскими белковыми молекулами
титина; молекулы миозина имеют длинный хвост и на его конце
две головки, при повышении концентрации ионов кальция в
области присоединения головок (в своеобразном шарнирном
участке) молекула миозина изменяет свою конфигурацию, при
этом (поскольку между миозиновыми филаментами
расположены актиновые) головки миозина связываются с
актином (при участии вспомогательных белков — тропомиозина
и тропонина), наклоняются и тянут за собой актиновую молекулу
в сторону М-линии, а их Z-линии сближаются и саркомер
укорачивается, при этом источником ионов кальция служат
цистерны агранулярной эндоплазматической сети, которые
вытянуты вдоль миофибрилл около каждого саркомера и
образуют саркоплазматическую сеть (в ней аккумулируются ионы
кальция, когда миосимпласт находится в расслабленном
состоянии); от поверхности миосимпласта плазмолемма образует
длинные трубочки, идущие поперечно в глубину клетки (Т-
трубочки) на уровне границ между темными и светлыми
дисками; когда клетка получает сигнал о начале сокращения, этот
сигнал перемещается по плазмолемме в виде потенциала
действия и распространяется отсюда на мембрану Т-трубочек и,
поскольку эта мембрана сближена с мембранами
саркоплазматической сети, состояние последних меняется,
кальций освобождается из цистерн сети и взаимодействует с
актино-миозиновыми комплексами, которые сокращаются, а
когда потенциал действия исчезает, кальций снова
аккумулируется в цистернах саркоплазматического ретикулума и
сокращение миофибрилл прекращается, но тогда для развития
усилия сокращения нужна энергия, которая и освобождается за
счет АТФ- АДФ-превращений (роль АТФазы выполняет миозин), а
источником АТФ служат митохондрии, поэтому они и
располагаются непосредственно между миофибриллами;
миоглобин и гликоген включён в деятельность миосимпластов
для выполнения следующих функций: гликоген служит
источником энергии, необходимой не только для совершения
мышечной работы, но и для поддержания теплового баланса
всего организма, а миоглобин связывает кислород, когда мышца
расслаблена и через мелкие кровеносные сосуды кровь может
свободно протекать, но при сокращении мышцы сосуды
сдавливаются (кровь не поступает), а запасенный кислород
освобождается из миоглобина и участвует в биохимических
реакциях.
Миофибри́ллы являются органеллами клеток
поперечнополосатых мышц и обеспечивают их сокращение
(служат для сокращений мышечных волокон), представляют из
себя нитевидную структуру, состоящую из структурных единиц
саркомеров, имеющих длину около 2 мкм и содержащих два
типа белковых филаментов: тонкие миофиламенты из актина и
толстые филаменты из миозина (именно их взаимодействие и
обеспечивает изменение длины мышечного волокна); границы
между саркомерами (Z-диски) состоят из особых белков, к
которым крепятся концы актиновых филаментов, а миозиновые
филаменты также крепятся к границам саркомера с помощью
нитей из белка титина (тайтина); с актиновыми филаментами
связаны также вспомогательные белки — небулин и белки
тропонин-тропомиозинового комплекса (тропонин, тропомиозин
и др); у человека толщина миофибрилл составляет 1—2 мкм, а их
длина может достигать длины всей клетки (до нескольких
сантиметров); одна клетка обычно содержит несколько десятков
миофибрилл и на их долю приходится до 2/3 сухой массы
мышечных клеток.
Миосателлитоцитами являются малодифференцированные
одноядерные клетки, являющиеся источником регенерации
мышечной ткани и прилежат к поверхности миосимпласта, таким
образом, что их плазмолеммы соприкасаются и обладают всеми
органеллами общего значения (в том числе и клеточным
центром); при повреждении мышечного волокна или длительном
увеличении нагрузки на мышцы клетки начинают делиться,
обеспечивая рост миосимпласта.
Скелетная мышца передаёт усилия своего сокращения на
костный скелет посредством сухожилий или прикрепления
непосредственно мышцы к надкостнице: на конце каждого
мышечного волокна плазмолемма образует глубокие узкие
выпячивания, в которые со стороны сухожилия или надкостницы
проникают тонкие коллагеновые волокна, спирально
сплетающиеся ретикулярными волокнами и концами
направленные к базальной мембране, войдя в нее, поворачивают
назад и при выходе снова оплетают коллагеновые волокна
соединительной ткани; между мышечными волокнами находятся
тонкие прослойки рыхлой волокнистой соединительной ткани
эндомизия, в который вплетаются коллагеновые волокна
наружного листка базальной мембраны, что способствует
объединению усилий при сокращении миосимпластов; несколько
мышечных волокон окружают более толстые прослойки рыхлой
соединительной ткани, образуя перимизий, разделяющий
мышцу на пучки, затем несколько пучков объединяются в более
крупные группы, разделенные более толстыми
соединительнотканными прослойками, а соединительная ткань,
окружающая поверхность всей мышцы, называется эпимизием.
Различные типы мышечных волокон в составе различных мышц
функционируют в неодинаковых биомеханических условиях и
поэтому обладают разной силой, скоростью и длительностью
сокращения, а также утомляемостью, ферменты в них обладают
разной активностью и представлены в различных изомерных
формах (различны в них содержания дыхательных ферментов —
гликолитических и окислительных); по соотношению
миофибрилл, митохондрий и миоглобина различают белые,
красные и промежуточные волокна, по функциональным
особенностям разделяются на быстрые, медленные и
промежуточные мышечные волокна и по организации миозина
различают на «быструю» и «медленную» изоформы.
Регенерация скелетной мышечной ткани (камбиальные
элементы) происходит за счёт миосателлитоцитов (ядра
миосимпластов делиться не могут, так как у них отсутствуют
клеточные центры), пока организм растет, они делятся, а
дочерние клетки встраиваются в концы симпластов и по
окончании роста размножение затухает, а после повреждения
мышечного волокна на некотором протяжении от места травмы
оно разрушается и его фрагменты фаго цитируются макрофагами.
Восстановление любых тканей организма может осуществляется
за счет двух механизмов: гипертрофии и гиперплазии: при
гипертрофии происходит компенсаторное увеличение объема
самого симпласта, за счет увеличения количества миофибрилл и
др., при этом в симпласте активизируются гранулярная
эндоплазматическая сеть и аппарат Гольджи и происходит синтез
веществ, необходимых для восстановления саркоплазмы и
миофибрилл, а также сборка мембран, так что восстанавливается
целостность плазмолеммы, при этом поврежденный конец
миосимпласта утолщается, образуя мышечную почку; при
гиперплазиии происходит пролиферация миосателлитоцитов и
сохранившиеся рядом с повреждением делятся: одни из них
мигрируют к мышечной почке и встраиваются в нее, другие
сливаются (так же, как миобласты при гистогенезе) и образуют
миотубы, которые затем входят в состав вновь образованных
мышечных волокон или формируют новые волокна.
Васкуляризацию (питание) мышц осуществляют артерии,
входящие в неё и распространяющиеся по прослойкам
соединительной ткани, постепенно истончаясь; ветви 5—6-го
порядка образуют в перимизии артериолы, а в эндомизии
расположены капилляры, которые идут вдоль мышечных
волокон, анастомозируя друг с другом. Венулы, вены и
лимфатические сосуды проходят рядом с приносящими
сосудами; рядом с сосудами много тканевых базофилов,
принимающих участие в регуляции проницаемости сосудистой
стенки.
Иннервация в мышцах осуществляется миелинизированными
эфферентными (двигательными), афферентными
(чувствительными), а также не миелинизированными
вегетативными нервными волокнами следующим образом:
отросток нервной клетки, приносящий импульс от моторного
нейрона спинного мозга, ветвится в перимизии, каждая его ветвь
проникает сквозь базальную мембрану и, у поверхности
симпласта на плазмолемме, образует терминали, участвующие в
организации так называемой моторной бляшки или нервно-
мышечного соединения; при поступлении нервного импульса из
терминали выделяется ацетилхолин — медиатор, который
вызывает возбуждающий потенциал действия,
распространяющееся отсюда по плазмолемме миосимпласта,
иннервируя каждое мышечное волокно самостоятельно, и,
окружая их сетью гемокапилляров, образует комплекс,
именуемый мионом (группа мышечных волокон, иннервируемых
одним мотонейроном, называется нервно-мышечной единицей,
а мышечные волокна, принадлежащие к одной нервно-
мышечной единице, расположены мозаично среди волокон,
относящихся к другим единицам); чувствительные нервные
окончания располагаются не на рабочих мышечных волокнах, а
связаны со специализированными мышечными волокнами в так
называемых мышечных веретенах, которые расположены в
перимизии; волокна в таких чувствительных мышечных
веретенах называются интрафузальными волокнами, а обычные
рабочие мышечные волокна – экстрафузальными.
Интрафузальные мышечные волокна веретен значительно
тоньше рабочих, и образуются из двух видов: волокон с ядерной
сумкой и волокон с ядерной цепочкой, при этом каждое
мышечное волокно веретена спирально обвито терминалью
чувствительного нервного волокна, в результате чего сокращения
или расслабления рабочих мышечных волокон изменяют
натяжение соединительнотканной капсулы веретена,
соответственно изменяя тонус интрафузальных мышечных
волокон, вследствие чего возбуждаются чувствительные нервные
окончания, обвивающие их, и в области терминалей возникают
афферентные нервные импульсы; на каждом миосимпласте
располагается также своя моторная бляшка, обуславливающая
постоянное нахождение в напряжении интрафузальных
мышечных волокон, подстраивающихся к длине мышечного
брюшка в целом.