Небольшое введение
Псевдозухии начали покорять планету ещё с конца раннего триаса, однако особых успехов достигли уже в конце среднего триаса, а точнее в ладинском ярусе, вытеснив архозаврообразных с ниш крупных хищников, а также вклинившись в ниши мелкоразмерных и среднеразмерных, где они конкурировали с цинодонтами.
В начале позднего триаса псевдозухии достигли своего пика, однако к его концу им пришлось уступить дорогу более продвинутым динозаврам.

Впрочем, обо всём по порядку...
Базальные псевдозухии
В позднем триасе всё ещё оставались базальные роды (например дасигнатоид) и даже целые семейства псевдозухий, такие как орнитозухиды.
Орнитозухиды
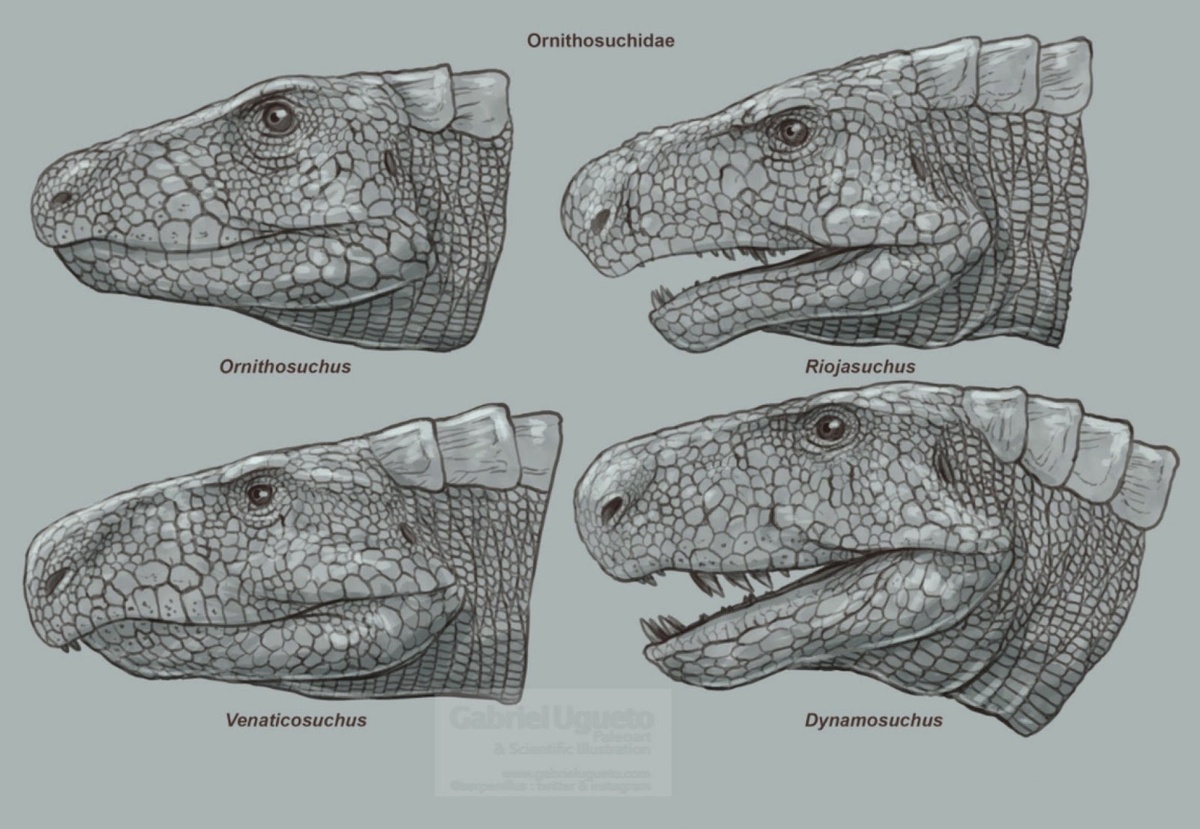
Орнитозухиды (лат. Ornithosuchidae, букв. «птицекрокодилы»), – вымершее семейство базальных псевдозухий, жившее на территории Лавразии (Великобритания) и Гондваны (Бразилия и Аргентина), в позднем триасе: 235-203,6 миллионов лет назад с карнийского по ретийский ярусы. В роду присутствует 4 подтверждённых вида и 1 спорный, таких как динамозух, орнитозух, венатикозух и риоджазух, ну а также энигмаспина, чьё положение в семействе является спорным.
Строение
Череп и зубы
Орнитозухид можно идентифицировать по наличию дугообразной диастемы , щели между зубами в передней части морды. Когда челюсть закрыта, два больших, изогнутых, зуба дентарии (нижней челюсти) входят в диастему, которая расположена между предчелюстной и верхней челюстью . На стенке диастемы есть два неглубоких углубления для размещения этих зубов. Большие зубы дентарии орнитозуха и риоджазуха расположены за меньшим, выступающим зубом дентарии, который торчит из челюсти. Такой тип расположения зубов не встречается ни у каких других базальных архозавров. Другой характерной чертой орнитозухид является их необычная загнутая вниз, нависающая морда, наблюдаемая у риоджазуха и венатикозуха , но не у орнитозуха.
Несколько других признаков отличают орнитозухидов от всех других ранних архозавров. У орнитозуха и риохазуха имеется небольшое отверстие (фенестра) между нёбными и крыловидными костями нёба , то есть нёбной кромкой. Контакт между носовыми и префронтальными костями черепа невелик или отсутствует, что исключается большим контактом между лобными и слёзными костями . У других архозавров, включая равизухий, этозавров, птерозавров и динозавроморфов, носовой и префронтальный контакт отделяет лобную кость от слёзной.
Посткраниальный скелет
Посткраниальный скелет почти полностью известен у риоджазуха, неполный у орнитозуха и совершенно неизвестен у венатикозуха. В результате, неизвестно, все ли посткраниальные черты, кажущиеся уникальными для орнитозухид, действительно наблюдались у всех членов семейства. Орнитозухиды, известные по достойным посткраниальным останкам, обычно имели около 9 шейных , 14-15 спинных, 3 крестцовых и более 20 хвостовых позвонков . Над каждым позвонком располагалась пара костных щитков, известных как остеодермы.
Бедренная кость (бедренная кость) имеет выраженный передний трохантер. Передний трохантер, иногда называемый «малым трохантером» (но не связанный с малым трохантером бедренной кости у человека), представляет собой гребень на наружной поверхности бедренной кости, рядом с головкой бедренной кости . Вероятно, он служил местом прикрепления m. iliofemoralis cranialis, которая помогает поднимать ногу. У большинства архозавров и их родственников выраженный передний трохантер отсутствует, но орнитозухиды, как и большинство динозавроморфов (динозавров и их близких родственников), являются исключением.
Подобно бедренной кости, малоберцовая кость (наружная часть большеберцовой кости) также имеет характерную точку прикрепления мышцы. Эта мышца называется подвздошно-офибулярной и помогает выпрямлять конечности. У большинства архозавров подвздошно-офибулярная мышца крепится к малоберцовой кости посредством крошечного гребня на проксимальной части малоберцовой кости, около колена. Однако у орнитозухид точка прикрепления подвздошно-офибулярной кости гораздо больше, имеет форму шишки и расположена примерно посередине диафиза малоберцовой кости. У фитозавров и этозавров также есть общая точка прикрепления в форме шишки, расположенная посередине малоберцовой кости, поэтому неясно, является ли случай орнитозухид уникальным случаем конвергентной эволюции или же сохранением признака, независимо утраченного несколькими линиями архозавров.
В отличие от большинства других ранних архозавров, когти стоп (дистальные кости стоп, образующие когти) сжаты с боков. Они острые и изогнутые. Когти очень глубокие, их высота превышает длину, особенно на внутренних пальцах. Этот тип когтей не встречается ни у одного другого триасового архозавра, за исключением птерозавров.
Основные группы архозавров часто различаются по строению голеностопных суставов. У большинства круротарзанов астрагал имеет выпуклый выступ, который входит в вогнутое пространство пяточной кости . Это состояние часто называют «крокодило-нормальным» голеностопным суставом, поскольку это наиболее распространённый тип голеностопного сустава у круротарзов. Орнитозухиды отличаются от круротарзанов и всех других архозавров наличием «крокодило-обратного» голеностопного сустава, в котором вогнутость расположена наоборот: вместо пяточной кости она расположена на астрагале. У орнитозухид пяточная кость имеет выпуклый выступ, аналогичный выпуклому выступу на «крокодило-нормальном» астрагале.
Образ жизни
Орнитозухиды были плотоядные, одни из них, такие как орнитозух, были апекс-хищниками в своей среде, другие, как динамозух и риоджазух были среднеразмерными хищниками, а венатикозух, например, был падальщиком.
Орнитозухиды были преимущественно четвероногими, однако они также имели адаптации к двуногости, что давало им преимущества над более медленными полностью четвероногими хищниками.
В ретийском ярусе позднего триаса орнитозухиды полностью вымирают, вероятно не выдержав конкуренции с набирающими силу динозаврами.
Зухии
Эрпетозухиды
Эрпетозухиды (Erpetosuchidae), также как в среднем триасе занимали ниши мелкоразмерных-среднеразмерных хищников.
В позднем триасе они представлены четырьмя родами, такими как пагосвенатор, эрпетозух, археопельта и диоплакс. Жили они на территориях восточной части США, Шотландии, Германии, а также Бразилии. Окончательно вымирают в конце норийского яруса ~208,5 миллионов лет назад.
Грацилизухиды
Грацилизухиды (Gracilisuchidae) как и в среднем триасе, были мелкоразмерными хищниками, охотившимися на насекомых и мелких тетрапод. В позднем триасе они представлены 3-мя родами, такими как: парвозух, грацилизух и спорный род маехари. Живут в основном на территориях Южной Америки.
Грацилизухиды вымирают в раннем норийском ярусе ~225,42 миллионов лет назад.
Этозаврообразные
Этозаврообразные (Aetosauriformes) - клада псевдозухий, которые впервые появляются в начале карнийского яруса ~237 миллионов лет назад. Этозаврообразных разделяют на базальных представителей и на, собственно, этозавров (Aetosauria).
Базальные этозавриформы, такие как ревуэлтозавр, эусколозух и аценазух, были зачастую мелкоразмерными всеядными животными, которые могли питаться как растительной, так и животной пищей.
У них уже была броня, которая использовалась для защиты их небольших тел. Некоторые из них, такие как ревуэлтозавр, возможно вели полуводный образ жизни.
Этозавры
Этозавры, или же Aetosauria ( / eɪˌɛt oʊˈsɔːr iə / ; от греч . ἀετός - aetos, «орёл», и σαυρος - sauros, «ящерица») – вымерший отряд бронированных псевдозухий, живших в позднетриасовую эпоху: 237-201,4 миллиона лет назад. Этозавры были весьма распространены на Пангее, они жили на территориях Европы, Северной и Южной Америк, а также в Северной Африке и Индии.
Классификация
Этозавры являются базальными представителями зухий. В состав отряда входит +20 родов, что говорит о весьма широком разнообразии. Подразделяются этозавры на три подсемейства: этозаврины, десматозухины и стагонолепидины, которые входят в состав одного подсемейства стагонолепид. В подсемействе этозаврин также присутствует клады типоторацин и паратипоторацин.
Описание
Череп
Череп этозавров относительно мал по сравнению с телом и имеет весьма характерную форму. Зубы отсутствуют как на передней части предчелюстной кости (кость, образующая кончик морды), так и на передней части зубной кости (зубчатая кость нижней челюсти).Присутствующие зубы обычно маленькие и выпуклые, от простых конических форм до листовидных с крупными зазубринами. Это, вероятно, указывает на всеядную или травоядную диету, и подобные адаптации наблюдаются у других архозавров, в меньшей степени полагающихся на мясо в своем рационе. У некоторых этозавров есть зубы зифодонтной формы, что означает, что зубы загнуты назад, зазубрены и уплощены сбоку. Эта форма, которая преобладает у этозавроида и небольшого образца, предположительно отнесенного к коахомазуху, типична для плотоядных архозавров.
У некоторых этозавров (отдельных членов группы десматозухий) кончик морды расширен вбок в уплощенную «лопатообразную» форму, похожую на морду свиньи . Внешние ноздри (ноздревые отверстия) удлиненные, намного больше, чем анторбитальные окна (отверстие на боковой стороне черепа). У многих этозавров есть небольшой выступ на предчелюстной кости, который выступает в ноздри снизу. У всех этозавров, за исключением этозавроида , задний край ноздрей получает вклад от вогнутого переднего края верхнечелюстной кости . В задней верхней части черепа отверстие, известное как надвисочное окно, расположено и выставлено на бок, в отличие от большинства других архозавров, у которых оно в основном видно при осмотре черепа сверху.Мозговая коробка довольно стандартна по стандартам псевдозухий, хотя отверстие для отводящего нерва проходит через парабазисфеноидную кость (в нижней передней части мозговой коробки), а не через предушную кость (в верхней передней части). Эта черта встречается только у ревуэльтозавра и крокодиломорфов среди архозавров.Нижняя челюсть (нижняя челюсть) описывается как имеющая форму «тапочки» у многих этозавров. Это связано с комбинацией особенностей: передняя часть зубной кости сильно сужается к точке, в то время как нижняя часть зубной кости иногда сгибается в «подбородок» (выступ вниз), который может также обнажать пластинчатую кость. Челюстной сустав расположен низко, а сочленовная ( кость нижней челюсти, которая соединяется с черепом) часто имеет высокий выступ прямо за челюстным суставом.
Посткраниал
Во многих отношениях, за исключением черепа и панциря, анатомия скелета этозавров была довольно стандартной среди других крупных триасовых псевдозухий. Задние конечности развили «столбовидную» позу, похожую на ту, что наблюдается у « рауизухий », родственного класса плотоядных триасовых псевдозухий, предков крокодиломорфов. Столбовидная поза конечностей - это та, при которой бедренная кость сочленяется вертикально с вертлужной впадиной бедра, которая наклонена вниз, так что нога располагается под телом и действует как опорная колонна. Тем не менее, вероятно, существовали значительные различия в строении тазобедренного сустава этозавров, и передние конечности, возможно, имели полураскинутую «гибридную» позу. Хотя строение задних конечностей похоже на равизухий, другие черты более плезиоморфны (типичны для предковых псевдозухий), такие как крепкий таз и широкие пятипалые ступни.Передние конечности были меньше задних, а лучевая кость , в частности, была намного короче плечевой. Тем не менее, их низкая и тяжелая форма тела требует, чтобы все этозавры были четвероногими . У них было несколько адаптаций для укрепления тела в ответ на их тяжелую броню: подвздошно-малоберцовая мышца крепилась в нижнем положении на малоберцовой кости , четвертый вертел бедренной кости был увеличен, поперечные отростки (крепления ребер) развились в длинные массивные пьедесталы, а самые крупные виды даже приобрели усиления гипосфена-гипантрума между позвонками.
Хотя этозавры в целом были широкотелыми рептилиями, существуют некоторые различия в степени выраженности этой тенденции. Типотораковые , представленные типотораксом и паратипотораксом , имели очень широкий, дисковидный панцирь, окаймленный небольшими шипами или килями и переходящий в узкий хвост. Самый крупный вид типотораксовых мог достигать около 3 метров в длину и 110 кг в весе. Десматозухины, такие как десматозух и лонгозух , имели умеренно более узкие тела и отсутствовали брюшные доспехи. Однако они также приобрели более шипастые доспехи на спине, особенно в шейном отделе (области шеи).Десматозух, вероятно, был одним из крупнейших известных этозавров, достигая 4–6 м в длину и веса 280-300 кг.
Этозавры, которые не попадают в эти две категории, такие как стагонолепис и неоэтозавроид , обычно имели узкие формы, тонкие конечности и ограничение в панцире выше бедра.Этот тип телосложения является плезиоморфным (предковым) по отношению к двум другим формам, при этом некоторые узкотелые аэтозавры более тесно связаны с типоторацинами, а другие ближе к десматозухинам. Некоторые плезиоморфные роды, такие как широко распространенный норийский род этозавров и карнийский род коахомазухов, имели тенденцию быть небольшими, около метра в длину. Другие были крупнее, такие как самый базальный этозавр этозавроид и ранний десматозухин калиптозух.
Броня
Этозавры были очень тяжело бронированы, с рядами больших, взаимосвязанных костных пластин, известных как остеодермы , которые защищали спину, бока, живот и хвост.Эти остеодермы обычно имеют четырехугольную (четырехстороннюю) форму и, несомненно, использовались в качестве защиты от хищников. Большинство остеодерм сильно изрыты на своих верхних поверхностях и гладкие на нижних. Они имеют гетерогенную внутреннюю структуру: внутренняя часть каждой остеодермы состоит из губчатой или губчатой кости (также называемой диплоэ), а их внешние части состоят из компактной кости. При жизни эти пластины, вероятно, были покрыты ороговевшим (роговым) покрытием, как современные крокодиловые щитки , которые являются еще одним примером псевдозухийных остеодерм.
Остеодермы полезны для диагностики таксонов этозавров, а виды этозавров часто можно идентифицировать по отдельным щиткам на основе их формы, структуры или рисунка орнамента.
Этозавры имеют четыре ряда остеодерм, идущих вдоль их дорсальной (задней) стороны, образуя непрерывную пластину, часто называемую карапаксом . Внутренние два ряда, которые обрамляют среднюю линию позвоночника, известны как парамедианные остеодермы. Они, как правило, шире, чем длиннее, и сильно украшены радиально расходящимися ямками или бороздками. Почти у всех этозавров есть небольшой бугорок или приподнятая поверхность, известная как дорсальное возвышение, на верхней поверхности каждой пластины. Дорсальное возвышение часто расположено сзади (назад) или медиально (внутрь) на их соответствующей парамедианной остеодерме, хотя внутри группы есть много исключений. Парамедианные остеодермы почти всегда имеют приподнятые или вдавленные передние края, где пластины перекрываются теми, что находятся перед ними. Если передний край приподнят, эта область называется передней полосой, а если он вдавлен, эта область называется передней пластиной. Хотя два парамедиальных ряда остеодерм часто встречаются у ранних архозавриформ, немногие рептилии приближаются по сложности строения к этозаврам. Остеодермы досвеллиид , эрпетозухид и некоторых крокодиломорфов иногда путают или сравнивают с таковыми этозавров. У аценазуха , близкого родственника этозавров, первоначально ошибочно идентифицированного как ювенильный десматозухин, встречаются как передняя перекладина, так и дорсальный выступ.
Внешние два ряда остеодерм, лежащие рядом с парамедианами, называются латеральными остеодермами. Они параллельны парамедианам почти по всей задней стороне, хотя первые две парамедианы позади головы (известные как затылочные остеодермы) являются одиночными. Боковые пластинки обычно разделены на две поверхности, или фланцы, согнутые между их дорсальными возвышениями. Верхний, или дорсальный фланец, лежит в той же плоскости, что и парамедианные остеодермы. Нижний/наружный, или латеральный фланец загибается на бок тела. Дорсальный выступ между этими выступами часто имеет форму невысокой лопасти, ручки или шипа. В шейных латеральных остеодермах, которые расположены на шее, дорсальный выступ имеет тенденцию проявляться в виде заметного шипа. Это явление доведено до крайности у десматозухов, таких как лонгозух и десматозуха, у которых шип увеличен в резко изогнутый рог.
У большинства этозавров (за исключением десматозухинов ) нижняя часть животного также защищена остеодермами. Эти вентральные (брюшные) остеодермы, как правило, меньше и плоские, чем дорсальные, и расположены в большее количество рядов (обычно 5-14 рядов), по крайней мере, в области туловища. Ряды вентральных остеодерм обычно изгибаются наружу и разделяются под бедром, оставляя широкий промежуток для клоакального отверстия. Вокруг этого отверстия у типоторакса, одного из немногих исключений из общего правила гладких вентральных остеодерм, встречаются крупные крючковидные шипы. Вентральные ряды распадаются на шагрень мелких пластин на шее и небольшое количество широких рядов под хвостом. Плотный набор мелких, не перекрывающихся пластин, известных как аппендикулярные остеодермы, покрывал передние и задние конечности.
Образ жизни
Этозавры в основном были растительноядными животными, хотя базальные их представители были всеядны. Большинство из них питалось мягкой растительностью у болот, а также корешками и клубнями, которые они выкапывали своими свиноподобными мордами. Возможно некоторые из этозавров (например лонгозух), были полуводными животными. Вымирают этозавры в конце ретийского яруса позднего триаса, тогда же, когда началось триасо-юрское вымирание.
Попозавроиды
Попозавроиды, попозавроидеи (Poposauroidea) или же «Группа X» в позднем триасе представлены двумя семействами – попозавридами (Poposauridae) и шувозавридами (Shuvosauridae). О них написано будет чуть ниже, а здесь рассмотрим особенности строения этой группы.
Череп и позвонки
Попозавроидов можно отличить от других псевдозухий по строению кончика морды, в частности, по предчелюстной кости , которая расположена перед ноздрями ( ноздрями ). Эта кость имеет два костных выроста («отростка»), охватывающих ноздри. Переднедорсальный отросток, который обхватывает ноздри и соприкасается с носовыми костями на верхнем крае морды, у псевдозухий обычно довольно короткий. У попозавроидов удлинённые переднедорсальные отростки, длиннее основного тела предчелюстной кости. Заднедорсальный отросток, который обхватывает ноздри и соприкасается с верхней челюстью по бокам морды, имеет противоположное строение. У попозавроидов он значительно короче (по сравнению с другими псевдозухиями), занимая лишь часть нижнего края ноздрей. Это позволяет верхней челюсти формировать остальную часть нижнего и заднего края ноздрей, при этом передний край верхней челюсти также становится вогнутым. Хотя эти особенности морды редки среди псевдозухий, они гораздо чаще встречаются у некоторых авеметатарзалий (архозавров птичьего ряда), таких как птерозавры и ящеротазовые динозавры . Задняя ветвь верхней челюсти сужается у большинства попозавроидов, за исключением квианозуха . Это контрастирует с лорикатами, у которых эта ветвь имеет прямоугольную форму.
Попозавроиды также обладают несколькими особенностями, которые необычны по сравнению с архозаврами в целом. Например, у большинства архозавров каждая сторона мозговой коробки имеет ямку, из которой внутренние сонные артерии могут выходить из мозга . У ранних попозавроидов эти ямки мигрировали на нижнюю сторону мозговой коробки, тем самым напоминая примитивное состояние, наблюдаемое у родственников архозавров, таких как эупаркерия и протерохампсии. Тем не менее, эта реверсия отменена у шувозавридов (и, возможно, раньше, хотя у попозавра или лотозавра материал мозговой коробки неизвестен). Кроме того, большинство попозавроидов обладали удлиненными шеями, и все они имели длинные и тонкие шейные ребра. Эта вторая черта шеи контрастирует с состоянием у других псевдозухий, фитозавров и птерозавров, у которых короткие и толстые шейные ребра. Остистые отростки спинных позвонков тонкие и пластинчатые, даже у представителей попозавроидов без парусов. Это отличает их от позвонков большинства других ранних псевдозухий (а также эупаркерии и фитозавров), у которых остистые отростки расширяются наружу, образуя при взгляде сверху плоскую прямоугольную поверхность.
Таз
Как и у других рептилий, таз попозавроидов образован тремя пластинчатыми костями: подвздошной костью , расположенной над вертлужной впадиной (тазовой впадиной), лобковой костью , расположенной ниже и спереди от вертлужной впадины, и седалищной костью , расположенной ниже и сзади от вертлужной впадины. Подвздошная кость — крупная, сложная кость с направленным вперёд (преацетабулярным) отростком, направленным назад (постацитабулярным) отростком и нижней частью, образующей верхний край вертлужной впадины. У большинства архозавров нижняя часть подвздошной кости имеет клиновидную форму, образуя внутреннюю поверхность «закрытой» вертлужной впадины. У попозавроидов нижняя часть подвздошной кости вогнута, что создаёт частично или полностью «открытую» вертлужную впадину, образованную открытым пространством вместо кости. Единственными другими архозаврами с открытыми вертлужными впадинами являются динозавры и (в меньшей степени) крокодиломорфы. Верхний край вертлужной впадины образован выраженным гребнем на подвздошной кости, известным как надвертлужный край или гребень.
Хотя все попозавроиды обладали открытой вертлужной впадиной, большинство других специализаций подвздошной кости не развивались до клады, содержащей попозавра и шувозавридов. Например, надвертлужный гребень в этой кладе выступает вниз, а не наружу. Эта черта редко встречается у архозавров, развиваясь независимо только у нескольких ранних тероподных динозавров, таких как целофиз и дилофозавр. Другой особенностью является наличие дополнительного диагонального гребня, который ответвляется вверх от надвертлужного края. Хотя такой гребень развился независимо у ряда различных архозавров, эта конкретная подгруппа попозавроидов уникальна тем, что гребень наклонен вперед (а не вертикально) и сливается с удлиненной преацетабулярной лопастью, что является еще одной производной чертой клады.
Лобковая и седалищная кости также были специализированы у попозавроидов. У всех остальных архозавров эти две кости соприкасались друг с другом на нижнем крае вертлужной впадины. У попозавроидов, за исключением квианозуха и лотозавра, кости не соприкасались, оставляя вертлужную впадину открытой с боков и снизу. Ширина лобковой кости варьируется в разных частях ее диафиза. Часть около вертлужной впадины утолщена, но кончик кости (за исключением квианозуха) очень тонкий, если смотреть спереди. У большинства других архозавров лобковая кость имеет постоянную ширину. У тероподовых динозавров и некоторых других архозавров дистальная часть лобковой кости тоньше проксимальной части. Шувозавриды и лотозавры также обладали седалищными костями (по обе стороны тела), которые были сращены друг с другом по средней линии тела.
Крестец
Попозавроиды имеют от трёх до четырёх крестцовых позвонков, причём последний и третий от конца позвонки сочленяются с подвздошной костью подобно двум первичным позвонкам более примитивных архозаврообразных, таких как эупаркерия и фитозавры. Предпоследний позвонок имеет форму, непохожую на позвонки этих архозаврообразных, и Несбитт пришёл к выводу, что это была «вставка», образованная из самых внутренних участков двух первичных позвонков. Хотя этот процесс не является уникальным для попозавроидов, он известен только в нескольких других группах архозавров, таких как батрахотомы , силезавриды и динозавры.
Базальные попозавроиды, такие как аризоназавр и квианозух, имели только три крестцовых позвонка, причём второй позвонок был «вставкой». Более продвинутые попозавроиды, такие как попозавр и шувозариды, имеют четыре крестцовых позвонка, третий из которых распознаётся как вставка. Это означает, что первый позвонок, должно быть, был ещё одним добавлением, по-видимому, последним спинным позвонком, который был перепрофилирован и преобразован в крестцовый позвонок. Этот позвонок включал спинной позвонок, называемый дорсо-сакральным. Они были неравномерно распространены среди архозавров, известны у некоторых орнитозухид и этозавров , а также у различных динозавров (чаще всего у птицетазовых и теропод).
Почти у всех архозариформ крестцовые ребра первого зачаточного крестцового позвонка контактируют с подвздошной костью около основания этой кости, близко к ее контакту с лобковой костью. Попозавроиды имели первые зачаточные крестцовые ребра с дополнительными передними ветвями, которые лежат на внутреннем крае преацетабулярной лопасти подвздошной кости. У попозавроидов, более продвинутых, чем квианозух, крестцовые позвонки срастаются в одну кость — крестец. Это слияние происходило постепенно, в разных частях позвонка. Например, зигапофизы срослись вместе уже у ктенозаврисцид. Центральные части (основная цилиндрическая часть) крестцовых позвонков также могли срастись уже у ктенозаврисцид. Основания невральных дуг (часть позвонков над спинным мозгом) были сращены у некоторых ктенозаврисков (аризоназавр), но не у других (брумсгрувия), а также были сращены у всех попозавроидов, более продвинутых, чем ктенозавриски.
Ещё некоторые особенности
В отличие от большинства псевдозухий, у попозавроидов отсутствуют костные щитки, известные как остеодермы . Единственным исключением является квианозух, у которого было множество крошечных остеодерм, лежащих в ряд и простирающихся вдоль шеи и тела. У всех попозавроидов кончик малоберцовой кости (внешняя берцовая кость) симметричный и прямой, если смотреть сбоку, а не наклонный, как у других некрокодиломорфных псевдозухий. У более продвинутых, чем ктенозаврисциды, были уплощенные копытообразные когти на ногах ( когти на пальцах ног). У некоторых попозавроидов были очень короткие руки по сравнению с длиной их ног, хотя экзартикуляция у квианозуха и отсутствие материала конечностей у ктенозаврисцид означает, что неизвестно, была ли эта черта базальной для группы в целом. Отсутствие данных по ктенозаврисцидам также не позволяет определить, когда определенные черты хвостовых позвонков и костей лодыжки были приобретены или утрачены в пределах попозавроидов.
Попозавриды
Попозавриды это семейство попозавроидов состоящее из двух родов: спорного долихобрахиума и валидного попозавра. Жили попозавриды на территориях Северной и Южной Америк, во времена позднего триаса: 237-216 миллионов лет назад. Это были бипедальные хищники среднего размера, которые охотились на разнообразную добычу включая как мелких так и средних животных. Вымерли попозавриды, скорее всего из-за конкуренции с динозаврами, хотя возможны и климатические причины.
Шувозавриды
Шувозавриды, другое семейство попозавроидов, но на этот раз растительноядных. Состоит это семейство из 3-х родов, таких как: силлозух, шувозавр и эффиджия. Жили шувозавриды с карнийского по ретийский ярусы: 235-201,3 миллиона лет назад, на территории Северной (США) и Южной (Аргентина) Америк. Шувозавр и эффиджия были средне/мелкоразмерными растительноядными, напоминавшими страусов или орнитомимид.
Силлозух же, возможно был крупноразмерным растительноядным животным, достигавшим 9,5-10 метров в длину, что делает его самым крупным из псевдозухий, за исключением разве что некоторых крокодиломорф.
Последним шувозавридом была эффиджия из Северной Америки, которая жила в ретийском ярусе позднего триаса и вымерла в результате массового вымирания в конце триасового и начале юрского периодов.
Лорикаты
Лорикаты были главными хищниками позднего триаса. Представлены они в основном двумя семействами – престозухидами и равизухидами. Престозухиды вымерли ещё в первой половине позднего триаса, тогда как вторые дотянули до позднего норийского яруса, правда вскоре вымерли из-за изменений климата. Самый крупный хищник триасового периода – фасолазух из норийского яруса Аргентины, был лорикатом и достигал длины в 10-12 метров, при весе до 1,6 тонны.
В Северной Америке обитал другой опасный хищник – постозух, напоминавший т-рекса из мелового периода. Он ходил на двух ногах, обладал хорошим зрением и нюхом, а его укус хорошо подходил для прокусывания панцирей или для дробления костей. Ещё одной важной особенностью постозуха была бипедальность, которая добавляла ему скорости.
Смерть псевдозухий и расцвет крокодиломорф
В конце карнийского яруса от одной из линий лорикат отделяются первые крокодиломорфы, те самые которые дадут начало современным крокодилам и аллигаторам. Большинство из них, как гесперозух, были мелкоразмерными хищниками охотившимися на насекомых и небольших животных. Правда некоторые, такие как карнифекс или редондавенатор смогли пробиться в средне- и даже крупногабаритный класс.
Крокодиломорфы будут единственной линией псевдозухий, которая доживёт до юрского периода и уже в нём начнётся новый расцвет этой группы.