Если вы палеогик 5-го уровня, вы не можете не знать об исключительно странной триасовой кладе дрепанозавроморфов (Drepanosauromorpha). Эти, как правило, маленькие рептилии с длинным телом, в основном, но не бесспорно, считаются находящимися у основания архозавроморф (что-то среднее между ящерицами и крокодилами) и известны своей крайне ненормальной анатомией. Изящные птичьи головы и шеи сидят на длинных, крепких и трубчатых телах с расширенными хвостами и крепкими конечностями. Конечности у каждого вида сильно видоизменены: у некоторых мощные когти, у других противопоставленные пальцы, как у хамелеона. Концы их хвостов преобразованы либо в хватательные органы, либо в острые крючки, что интерпретируется как приспособление для закрепления хвоста на растительности или субстрате. Образ жизни дрепанозавров долгое время был предметом обсуждения среди ученых, и в настоящее время их обычно считают древесными или роющими животными — или смесью того и другого. Они довольно удивительные создания.

Поскольку триасовый период был подпитываемым эволюцией бунтарскими студенческими днями, нельзя считать шокирующим узнать, что существует вид дрепанозавров, который для этой группы является тем же, чем они являются для всего остального. Этот уникальный, странный и противоречивый вид — Hypuronector limnaios. Достаточно хорошие окаменелости этого небольшого (около 12 см в длину) животного находили в течение десятилетий из верхнетриасовых отложений Нью-Джерси, но свое название оно получило сравнительно недавно (Colbert and Olsen 2001). Гипюронектор часто рассматривается как плавающее существо из-за его расширенного в дорсовентральном направлении «листообразного» хвоста, на котором отсутствует крючковатый или цепкий конец (Colbert and Olsen 2001). Его хвост примечателен огромными шевронами (зубцами кости, выступающими вниз с нижней стороны хвостовых позвонков), которые простираются далеко ниже и позади исходного позвонка, создавая большую часть глубины хвоста и его «листовидный» профиль. Некоторые авторы сравнили очертания хвостового скелета с формой тела гимнтоидной или гимнархидной рыбы и предположили, что она продвигала гипюронектора через глубокие пресноводные озера, в которых были захоронены его окаменелости, возможно, подобно тритону или крокодилу. Хотя они обладали необычно длинными ногами по сравнению с другими дрепанозаврами и плавающими животными, утверждалось, что они также были связаны с водным образом жизни. В частности, предполагается, что они держали длинный широкий хвост над землей во время наземных схваток, причем хвост, по-видимому, не мог подняться у основания (Colbert and Olsen 2001). Эта водная гипотеза существует уже некоторое время. В 1980-х годах это животное было неофициально известно как «глубокий хвостатый пловец» (Fraser and Renesto 2005), а в 2001 году это прозвище было более или менее полностью перенесено в его научное название: в вольном переводе Hypuronector означает «широкохвостый озерный пловец».
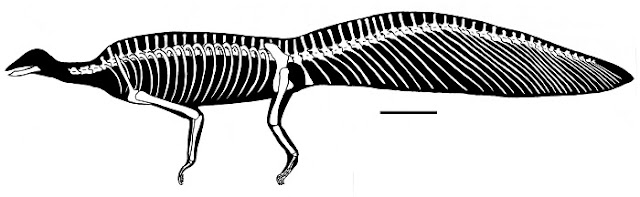
По крайней мере, на первый взгляд, все это не кажется слишком диковинным: хвост гипюронектора определенно имеет веслообразную форму, и все мы знаем, что боковые изгибы хвоста — самый распространенный способ передвижения по воде у позвоночных. Но есть и другие интерпретации гипюронектора, которые предполагают, что он вообще не был пловцом. Эти альтернативные точки зрения предполагают, что он больше походил на других дрепанозавров в плане приспособленности к лазанию, но, что более примечательно, возможно, он был планером (Renesto et al. 2010). Обмен ранними версиями моего рисунка планирующего дрепанозавра (выше) предполагает, что последняя гипотеза не очень известна даже среди экспертов. Однако сразу хочу подчеркнуть, что это не художественная спекуляция в духе All Yesterdays или причудливое мнение «маргинального» рабочего. Некоторые авторы (например, Senter 2004; Spielmann et al. 2006; Renesto et al. 2010; Castielloa et al. 2015) оспаривают концепцию водного гипюронектора и предполагают, что он был более «типичной» древесной формой. Предположение о том, что это мог быть планер, было выдвинуто на основании разумных (хотя, возможно, еще не убедительных) доказательств (Renesto et al. 2010). Это следует из более старых предположений о том, что некоторые дрепанозавриды, особенно мегаланкозавры (Megalancosaurus Calzavara, Muscio & Wild 1981), были планерами (см. ниже; Ruben 1998; Renesto 2000), и, хотя все это может показаться странным, для этого есть некоторая подлинная научная основа.
Водная гипотеза под лупой
Водная гипотеза была впервые предложена в начале 90-х годов (Berman and Reisz 1992) и быстро подверглась критике со стороны исследователей дрепанозавров (историю см. в Renesto 2010). Hypuronector, возможно, остается лучшим кандидатом на роль водного или, по крайней мере, земноводного вида из-за его необычного хвоста, но по иронии судьбы именно этот веслообразный орган кажется главной проблемой для этой гипотезы.
Одна вещь, на которую мы должны обратить внимание, заключается в том, что сходство хвоста гипюронектора с телом некоторых рыб не является лучшим подтверждением привычек плавания. Рыбы не плавают, используя все свое тело (передняя часть любого волнообразного пловца должна быть жесткой), а гимнтоидные или гимнархидные рыбы, сравниваемые с хвостом гипуронектора, на самом деле вообще не двигают своим телом во время плавания. Скорее, они движутся за счет колебаний длинных низких плавников вдоль верхней и нижней частей тела. Таким образом, они могут быть плохим аналогом органа для гребли, и нам лучше смотреть на плавники и ласты плавающих животных, а не на все их тела, чтобы узнать о водном потенциале хвоста гипуронектора.
Само собой разумеется, что гипюронектор плавал бы, как крокодил, тритон или плавающая ящерица, то есть волнообразные движения хвоста создавали бы толчок вперед (Colbert and Olsen 2001). Для этого требуется анатомия хвоста, которая может приспособиться к большому количеству боковых движений, и именно здесь Renesto et al. (2010) предполагают, что мы столкнулись с серьезной проблемой. Хвостовые позвонки гипюронектора, по-видимому, допускал некоторые движения в основании и на кончике хвоста, но середина хвоста была довольно жесткой. Это связано с тем, что зигаопофизы — костные отростки, перекрывающие соседние позвонки для направления их движения, — очень длинные и имеют крутые суставные поверхности (ниже). Проще говоря, они, кажется, «зажали» соседние позвонки, а не, — как и ожидалось от пловца с веслообразным хвостом, — обеспечили плоские горизонтальные поверхности, по которым позвонки могли бы скользить.
Дополнительная жесткость обеспечивается этими удивительными шевронами (Renesto et al. 2010). Эти выступающие назад кости лежат в основе сочленений соседних 7–8 позвонков, а это означает, что любое боковое движение в позвоночных суставах должно преодолевать жесткость 7–8 костных стержней, свисающих под ними. Хотя тонкие кости в некоторой степени податливы, а шевроны гипюронектора, возможно, были до некоторой степени гибкими, трудно представить, что их расположение оптимизировано для привычек гребли: они, возможно, сделали хвост более веслообразным, но с очевидным ущербом гибкости хвоста и потенциалу гребли. Действительно, мы должны отметить, что эта конфигурация очень похожа на биологические структуры, приспособленные к сопротивлению изгибу. Крылья четвероногих являются хорошим примером: расположение пальцев летучих мышей, структурных волокон птерозавров и стержней птичьих перьев по отношению к костям крыльев перекликается с распределением шевронов у гипюронектора. Напротив, у широкохвостых пловцов, таких как крокодилы и тритоны, шевроны короткие, крепкие и не сильно лежат под соседними позвонками. Они идеально подходят для закрепления мускулатуры хвоста, увеличения ширину хвоста и не мешают движению хвоста. Я должен согласиться с Renesto et al. (2010), что потенциал хвоста гипюронектора как плавательного органа кажется ограниченным.
Еще большее значение здесь имеют конечности гипюронектора, которые не имеют очевидных водных адаптаций. Водные или даже полуводные животные, как правило, имеют пропорционально короткие приземистые конечности, часто с расширенными лопатообразными костями. Но конечности гипюронектора удлиненные, изящные и полые (Renesto et al. 2010). Его передние и задние конечносты известны по неполному материалу и интерпретируются по-разному, но имеющиеся у нас элементы предполагают, что они не были похожи на весла. Колберт и Олсен (2001) предположили, что конечности гипюронектора были длинными, чтобы поднимать хвост от земли, когда он покидал воду, и их работа предполагала, что позвоночник был слишком жестким, чтобы поднимать хвост самостоятельно. Но это может показаться проблематичным по трем причинам. Во-первых, как указали Renesto et al. (2010), сочлененные окаменелости гипюронектора показывают, что хвост изгибается дугой вверх по отношению к позвонкам туловища (вверху): это не считается тафономическим или диагенетическим искажением. Во-вторых, передние конечности гипюронектора несколько длиннее задних, что, возможно, противоположно тому, что мы ожидали бы, если бы волочение хвоста вызывало беспокойство — наверняка тело наклонялось бы назад при таком расположении? В-третьих, с каких это пор рептилиям, водным или иным, стало плевать на хвосты? Мы должны быть осторожны, чтобы не оказывать «пустую поддержку» гипотезам, придумывая проблемы, которые должны решать наши ископаемые животные.
Может быть, Гипюронектор все-таки не «странный дрепанозавр»?
Взятые вместе, эти аспекты формы хвоста, его артрологии и размера конечностей должны рассматриваться как проблематичные для водной гипотезы, и, возможно, нам следует посмотреть, существуют ли другие интерпретации образа жизни гипюронектора, которые больше соответствуют его анатомии. Хорошая стратегия для понимания странных ископаемых животных состоит в том, чтобы отложить спорные, странные части анатомии в сторону и сначала сосредоточиться на более надежно интерпретируемых компонентах. С учетом сказанного, давайте на мгновение проигнорируем спорный хвост гипюронектора и посмотрим на его конечности, пояса конечностей и анатомию туловища. Как и у всех дрепанозавров, плечевые и тазовые кости гипюронектора очень высокие и чем-то напоминают пояса конечностей хамелеонов (Renesto et al. 2010). Считается, что оба набора конечностей были очень подвижными, хотя слияние грудного пояса дрепанозавроморфов в одну твердую структуру, в отличие от наличия двух отдельных половин, как у хамелеонов, несколько ограничило бы досягаемость передних конечностей. Конечности, вероятно, находились в растянутой позе, и, поскольку бедра и плечевые кости сильно удлинены, гипюронектор, вероятно, имел широкое и устойчивое основание, на котором можно было ходить и стоять.
Гипюронектор не имеет больших сросшихся позвонков над грудной областью, которые мы видим у других дрепанозавроморфов, но, учитывая, что они, вероятно, отражают увеличенную мышечную массу передних конечностей и усиленную грудную область для копания и захвата добычи (Castielloa et al. 2015), это может быть не так. Слишком сильно это влияет на двигательную механику. Туловище гипуронектора, очевидно, все равно имело мощную мускулатуру: высокие невральные шипы спинных позвонков и наличие больших изогнутых ребер вдоль всего туловища предполагали, что большая часть тела была покрыта крупными мышцами.
Можно видеть, что анатомия туловища и конечностей гипюронектора очень хорошо соответствует тому, что мы видим у других дрепанозавров: мощное туловище и подвижные конечности, которые, кажется, хорошо приспособлены для ходьбы и лазания. Мы могли бы рассматривать удлинение его конечностей как адаптацию к лазанию, увеличение длины сегментов верхних конечностей одновременно увеличивает стабильность и увеличивает досягаемость, а также удерживает центр масс близко к субстрату. Что еще более удивительно, гипюронектор также похож на других дрепанозавров в некоторых аспектах анатомии хвоста. Хотя его хвост имеет другую общую форму и не имеет производных от настоящих дрепанозавров кончиков хвоста, он разделяет особенности движения хвоста дрепанозавров - гибкое основание и кончик, жесткая середина - с остальной частью группы (Renesto et al. 2010). Так что, возможно, хвост гипюронектора был просто более простым вариантом хвоста дрепанозавроморфа странной формы и использовался для тех же целей: устойчивость при лазании (простая опора может помочь тяге, балансу и восстановлению после аварии), скоба при подъеме на дыбы для копания или просто для привлечения внимания (Renesto et al. 2010).
Сопоставив эти доказательства вместе, несколько авторов начали интерпретировать гипюронектора как более «типичного» дрепанозавра, хотя и менее специализированного вида, который жил как современная древесная ящерица, а не как древесный панголин или карликовый муравьед (Spielmann et al. 2006; Renesto et al. 2010). Если это правда, мы могли бы рассматривать форму его хвоста как механический отвлекающий маневр, что кажется более важным для поведения Гипуронектора, чем было на самом деле. Возможно, они имели не большее значение для передвижения и поведения, чем черепные украшения динозавров и птерозавров, структуры, которые, по мнению большинства, больше связаны с коммуникацией и демонстрацией, чем с механикой повседневной жизни.
Но что насчёт планирования?
Развивая эту идею древолазания, универсал гипюронектор сделал еще один шаг вперед. Группа Ренесто отметила, что у него есть несколько особенностей, которые могут указывать на то, что это был патагиумный планер, то есть животное с перепонками, находящимися между его конечностями, чтобы облегчить более медленное падение с возвышенности или парение между опорами. Главные особенности, представляющие интерес здесь, - это удлиненные конечности и, в частности, передние конечности такой же длины, если не чуть большей, чем задние. Эта конфигурация необычна среди рептилий. Хорошо известные рептилии с непропорционально длинными передними конечностями включают травоядных динозавров, просматривающих полог, полностью водные линии, такие как ихтиозавры, производные завроптеригии и черепахи, а также летающих животных, вроде птерозавров. Понятно, что эти животные ведут совершенно ой образ жизни, чем дрепанозавры, но основное сходство между тогдашними птерозаврами — мелких грацильных форм, у которых, вероятно, были лазающие и планирующие предки — и гипюронектором может быть немного более интригующим. Удлинение передних конечностей происходит снова и снова у патагиально скользящих четвероногих — птерозавров, шерстокрылов, чешуйчатохвостых планеров и т. д. — и вполне разумно задаться вопросом, выдает ли то же самое явление у гипюронектора наличие скользящих перепонок. Пропорции конечностей этого вида не настолько экстремальны, чтобы думать, что это был образцовый планер, способный преодолевать большие расстояния с вертикального старта, но они могли иметь перепонки достаточного размера, чтобы смягчить падение этих маленьких животных, если они прыгали или падали с высоких мест. Здесь также следует обратить внимание на широкую округлую форму хвоста, возможно, он имеет правильную форму, чтобы помочь «поставить» кувыркающегося гипюронектора в правильную позу для устойчивого планирования.
Как отмечалось выше, по крайней мере, мегаланкозавр также рассматривался как потенциальный планер в прошлом (Ruben 1998; Renesto 2000). Эти разговоры были вдохновлены (по крайней мере, частично) давно исчезнувшими (если вы когда-либо могли считать их заслуживающими доверия!) идеями о том, что у птиц могли быть общие предки с дрепанозаврами или дрепанозавроподобными животными — давайте быстро отойдём от дальнейшего обсуждения этого. Но почему идея планирующего мегаланкозавра не прижилась? Хотя это и не исключено полностью (Renesto 2000), планирование, похоже, не прижилось у этого вида, потому что его хвост с шипами, очень подвижные запястья и лодыжки, а также хватательные придатки предполагают, что он был весьма приспособлен к лазанию. Хотя лазание и скольжение не являются несовместимыми, ему также не хватает таких особенностей, как длинные изящные конечности, которые мы ожидаем от летающих животных. Анатомия гипюронектора, напротив, немного более обобщена и ставит достаточно галочек в колонке планеров, чтобы думать, что это возможно.
Конечно, стоит подчеркнуть, что любой планирующий дрепанозавр на данном этапе является гипотетическим, но мы не должны воспринимать это как причину, чтобы отвергать данную идею с ходу. В дополнение к свидетельствам, упомянутым выше, примите во внимание, что многие, а возможно, и все дрепанозавроморфы, по-видимому, были лазающими животными того или иного вида, и мы знаем из существующих фаун, что шаг от лазания к планированию часто бывает коротким (Renesto 2000). На самом деле глупо представить, что вымершие линии были менее способны развивать планирующие формы, чем наши современные, и дрепанозавры были приспособлены к планирующему полету во многих отношениях. У их черепов был большой мозг и перекрывающиеся поля зрения (Renesto and Dalla Vecchia 2005) (идеально подходит для оценки расстояния и обработки полетных данных); как правило, это были мелкие животные с полыми костями конечностей (легкие); их туловища были жесткими и укреплёнными (содействуют стабильности); их конечности были очень мускулистыми и подвижными (управление аэродинамическими крыльями), а их широкие, сильные хвосты могли быть идеальными рулями направления и стабилизаторами. И каким бы странным ни казалось обсуждение возможности планировать у животных, известных только по костям, вспомним, что птерозавры были идентифицированы как летающие животные в начале 1800-х годов, задолго до того, как мы обнаружили ископаемые остатки перепонок их крыльев: мы можем идентифицировать летающих животных, если мы достаточно внимательно посмотрим на их кости. Теперь задача состоит в том, чтобы проверить, сможем ли мы проверить эти идеи, возможно, тщательно сравнив анатомию конечностей и миологические признаки гипюронектора с другими дрепанозаврами, моделируя эффекты, которые ненормальный хвост оказывает на падающее животное, и так далее. Мы также можем искать перепонки на окаменелостях гипюронектора, исследуя их в ультрафиолетовом свете и проявляя особую осторожность при подготовке будущих образцов этого дрепанозавра: опыт с другими тонкими образцами рептилий показывает, что это помогает знать, где ожидать мягких тканей при снятии матрицы.
Итак, вот и все: триас и дрепанозавры, возможно, стали еще более странными/крутыми/сложными/разочаровывающими, чем мы все знали. Я думаю, нам нужно еще больше поговорить о триасовом периоде в будущих сообщениях в блоге — проверьте этот ярлык для предыдущих разговоров на триасовые темы. И обратите внимание, что в моей новой книге по искусству «Воссоздание эпохи рептилий» есть несколько страниц, посвященных триасовым животным, включая дрепанозавра.
Примечание от 07/06/2016:
Благодаря Андреа Кау, ниже было сделано несколько дополнительных цитат и вопросов для обсуждения - суть и аргументы поста те же, но контекст улучшен. Спасибо Андреа!
Марк Уиттон, 2016 год
Источник
См. также
#интересные факты #научпоп #рептилии #животные #биология #палеонтология