Основы редокс реакций.
- Таким образом, как и в случае с Ca2+, плейотропный характер окислителей влияет одновременно на множество фундаментальных процессов, что приводит к широко распространенным последствиям для здоровья и заболеваний.
- Santolini, J., Wootton, S. A., Jackson, A. A. & Feelisch, M. The redox architecture of physiolog
- В этом обзоре мы рассматриваем генерацию АФК и ее регулирование с акцентом на H2O2 и O2 ·-, их механизмы передачи окислительно-восстановительных сигналов, основные молекулярно-физиологические мишени окислительно-восстановительного потенциала и связанные с ними клеточные реакции на стресс.
- Мы также обсуждаем клеточные функции и процессы, на которые влияет АФК, а также дерегуляцию окислительно-восстановительной сигнализации при старении и некоторых патологических состояниях, с перспективой Перспективы окислительно-восстановительной медицины.
- Генерация и регуляция АФК АФК генерируются из различных источников. Чтобы сохранить свои уровни в физиологических концентрациях, различные механизмы контроля продукции и доступности АФК, включая локализованную и разделенную генерацию а также вовлечение поглотителей (детоксицирующих факторов) и окислительно-восстановительные реле установлены (рис. 2).
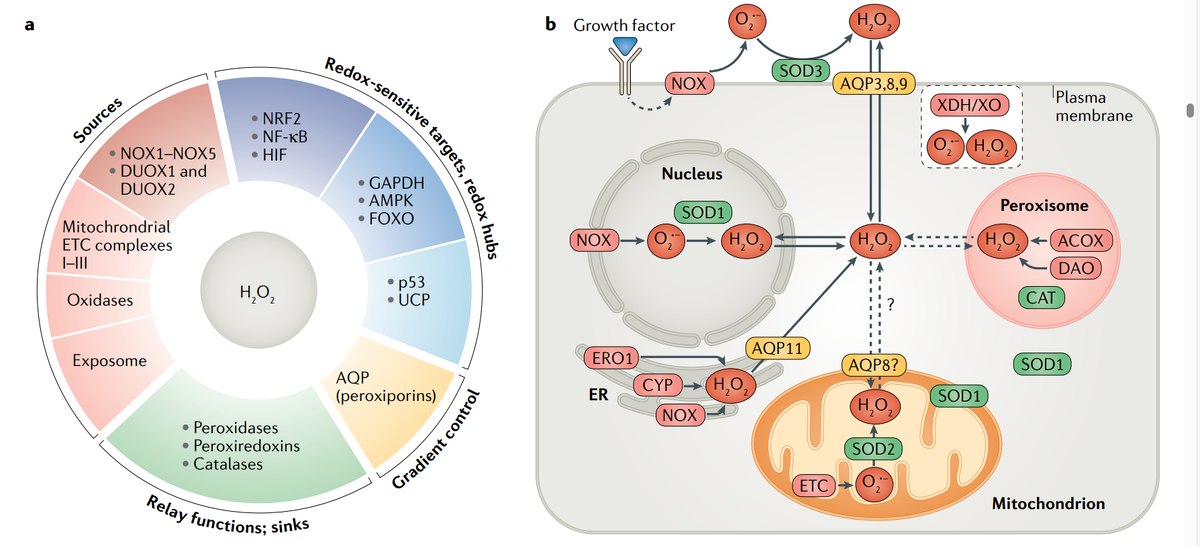
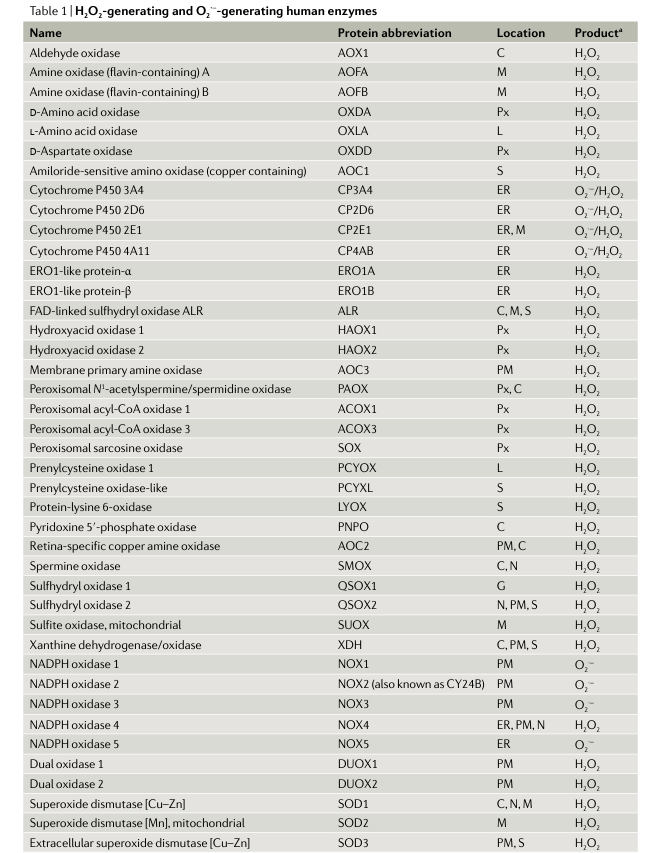
- Эндогенные и экзогенные источники АФК. В целом, в клетки человека всего 41 H2O2- и/или O2 ·--генерация ферменты были идентифицированы42 (табл. 1), и этот список увеличивается до более чем 50 за счет включения ферментов, генерирующих другие АФК, такие как гидроперекиси липидов или азотная кислота. оксид (NO) и хлорноватистой кислоты (дополнительный Таблица 1).
- Основные эндогенные ферментативные источники О2 ·- и H2O2 представляют собой трансмембранные НАДФН-оксидазы. (NOXs)и митохондриальная цепь переноса электронов (и т. д.).
Bedard, K. & Krause, K. H. The NOX family of ROS- generating NADPH oxidases: physiology and pathophysiology. Physiol. Rev. 87, 245–313 (2007).
Knock, G. NADPH oxidase in the vasculature: expression, regulation and signalling pathways; role in normal cardiovascular physiology and its dysregulation in hypertension. Free Radic. Biol. Med. 145, 385–427 (2019).
Parascandolo, A. & Laukkanen, M. O. Carcinogenesis and reactive oxygen species signaling: interaction of the NADPH oxidase NOX1-5 and superoxide dismutase 1-3 signal transduction pathways. Antioxid. Redox Signal. 30, 443–486 (2019).
Murphy, M. P. How mitochondria produce reactive oxygen species. Biochem. J. 417, 1–13 (2009). A comprehensive account of mitochondrial ROS production.
- NOX занимают различные клеточные локализации. (табл. 1), что способствует локальной генерации РОС. NOX также связаны со специализированными редокс-активные эндосомы (редоксосомы), образующие в ответ на специфические внеклеточные стимулы, такие как питательные вещества, факторы роста и цитокины, а также позволяют компартментализировать H2O2 для локальных окислительно-восстановительных процессов. регуляция (микродомены) или передача сигналов клетки от рецепторы клеточной поверхности47,48. В митохондриальной ETC, комплекс I и частично комплекс II выделяют O2 ·-/H2O2 по направлению к митохондриальному матриксу, тогда как высвобождение от комплекса III – в сторону просвета крист и межмембранное пространство.
Brand, M. D. Mitochondrial generation of superoxide and hydrogen peroxide as the source of mitochondrial redox signaling. Free Radic. Biol. Med. 100, 14–31 (2016).
- Это топологическое различие имеет функциональное значение, которое проявляется в различиях в структурах редокс-модифицированных белков в зависимости от источник.
Bleier, L. et al. Generator- specific targets of mitochondrial reactive oxygen species. Free Radic. Biol. Med. 78, 1–10 (2015).
- Помимо NOX и ETC, H2O2 также генерируются различными другими оксидазами, присутствующими в субклеточных локализациях, в основном включая эндоплазматический ретикулум (ЭР) и пероксисомы, а также несколько супероксиддисмутаз (СОД1–СОД3) (табл. 1), которые способствуют локализованному производству H2O2 из О2 ·− (рис. 2б). Стоит отметить, что помимо биология O2 ·-/H2O2, важная область исследований АФК касается АФК липидного происхождения, при этом полиненасыщенные жирные кислоты окисляются, образуя гидроперекиси липидов. и родственные радикалы, пероксил и алкоксил (дополнительные Таблица 1). Такие окисленные липиды оказывают большое влияние на Передача окислительно-восстановительных сигналов, особенно передача иммунных сигналов.
Higdon, A., Diers, A. R., Oh, J. Y., Landar, A. & Darley- Usmar, V. M. Cell signalling by reactive lipid species: new concepts and molecular mechanisms. Biochem. J. 442, 453–464 (2012).
Hitzel, J. et al. Oxidized phospholipids regulate amino acid metabolism through MTHFD2 to facilitate nucleotide release in endothelial cells. Nat. Commun. 9, 2292–04602 (2018).
Kagan, V. E. et al. Redox phospholipidomics of enzymatically generated oxygenated phospholipids as specific signals of programmed cell death. Free Radic. Biol. Med. 147, 231–241 (2020).
Spickett, C. M. & Pitt, A. R. Oxidative lipidomics coming of age: advances in analysis of oxidized phospholipids in physiology and pathology. Antioxid. Redox Signal. 22, 1646–1666 (2015).
Tyurina, Y. Y. et al. “Only a life lived for others is worth living”: redox signaling by oxygenated phospholipids in cell fate decisions. Antioxid. Redox Signal. 29, 1333–1358 (2018).
Эндогенные и экзогенные источники АФК. В целом, в клетках человека идентифицировано 41 фермент, генерирующий H2 O2 и/или O2 ·-42 (таблица 1), и этот список увеличивается до более чем 50 за счет включения ферментов, генерирующих другие АФК, такие как гидропероксиды липидов или азотистые кислоты, оксид (NO) и хлорноватистая кислота (дополнительная таблица 1). Основными эндогенными ферментативными источниками O2 ·- и H2 O2 являются трансмембранные НАДФН-оксидазы (NOX)43–45 и митохондриальная цепь переноса электронов (ETC)46. NOX занимают различные клеточные локализации (табл. 1), что способствует локальной генерации АФК.
NOX также связаны со специализированными окислительно-восстановительными активными эндосомами (редоксосомами), которые образуются в ответ на специфические внеклеточные стимулы, такие как питательные вещества, факторы роста и цитокины, и позволяют компартментализировать H2 O2 для локальной окислительно-восстановительной регуляции (микродомены) или передачи клеточных сигналов. от рецепторов клеточной поверхности.
Spencer, N. Y. & Engelhardt, J. F. The basic biology of redoxosomes in cytokine- mediated signal transduction and implications for disease- specific therapies. Biochemistry 53, 1551–1564 (2014)
. Mishina, N. M. et al. Imaging H2 O2 microdomains in receptor tyrosine kinases signaling. Methods Enzymol. 526, 175–187 (2013).
- В митохондриальной ЭТС комплекс I и частично комплекс II высвобождают O2 ·-/H2 O2 в направлении митохондриального матрикса, тогда как высвобождение из комплекса III происходит в направлении просвета крист и межмембранного пространства. Это топологическое различие имеет функциональное значение, о чем свидетельствуют различия в структуре редокс-модифицированных белков в зависимости от источника.
Давний вопрос заключается в природе преобладающих внутриклеточных генераторов оксидантов. Недавняя оценка относительного вклада NOX и митохондриальных сайтов ETC в покоящиеся миобласты показала, что Pr-(Tyr)-NO2 SSH N H O Персульфид H2 S SSR N H O Дисульфид SO–3 N H O Сульфонатный вклад различных источников в клеточный пул АФК зависит от контекст данной клетки и данного метаболического состояния и, следовательно, действительно различается. также источники o2 ·– и его продукта H2 o2 . Кристы складки внутренней мембраны митохондрий, что около 40% чистого производства H2O2 клетками приходится на NOX и примерно 45% приходится на внеземные земные шары, а остальная часть поступает из других ферментативных источников.
Wong, H. S., Benoit, B. & Brand, M. D. Mitochondrial and cytosolic sources of hydrogen peroxide in resting C2C12 myoblasts. Free Radic. Biol. Med. 130, 140–150 (2019).
Таким образом, в данном случае вклад NOX и ETC соизмерим. Тем не менее, помимо внутриклеточных источников, оксиданты также образуются в результате кумулятивного воздействия на окружающую среду, называемого «экспосомой», которое включает молекулярные факторы, такие как питательные вещества, лекарства, токсиканты и загрязнители, а также физические стрессоры (УФ-излучение). , рентгеновское излучение и другое ионизирующее излучение) и психологические стрессоры (Стиль жизни). Поскольку эти воздействия сильно различаются, трудно определить вклад экспосома в общий пул окислителей.
Управление H2O2 через поглотители и окислительно-восстановительном реле.
Пероксиредоксины и глутатионпероксидазы катализируют удаление H2O2. Эти ферменты имеют высокие константы скорости второго порядка в реакции с H2O2, порядка 105–108 М-1 с-1, что способствует поддержанию низкой клеточной концентрации H2 O2 в субклеточных пространствах, где они расположены15. Каталаза, гем-белок, дисмутирует H2 O2 до H2 O и O2 в каталатической реакции, а в пероксидатической реакции восстанавливает H2 O2 до H2 O путем окисления соединений, отдающих водород24. Таким образом, эти ферменты являются поглотителями H2O2. Некоторые пероксидазы, такие как миелопероксидаза, используют H2 O2 для генерации других оксидантов, таких как хлорноватистой кислоты, которая используется нейтрофилами для защиты от патогенов61, тем самым переназначая АФК для иммунной функции. Пероксиредоксины способны передавать окислительные эквиваленты от H2 O2 к другим белкам-мишеням, константы скорости реакции с H2 O2 которых сравнительно невелики. Примеры такого окислительно-восстановительного реле были продемонстрированы для пероксиредоксина 2 и транскрипционного фактора STAT3 (reF.30), а также для киназы в стресс-чувствительном сигнальном пути p3862.
Митохондриальная никотинамиднуклеотидтрансгидрогеназа (NNT) также играет роль в очистке клеточного (в том числе экстрамитохондриального) фактора роста b H2 O2 (reF.63). Это происходит за счет смещения восстанавливающих эквивалентов от НАДН к НАДФН, тем самым поддерживая и усиливая мощность тиоредоксиновой системы и системы глутатиона, которые зависят от поступления НАДФН.
Hanschmann, E. M., Godoy, J. R., Berndt, C., Hudemann, C. & Lillig, C. H. Thioredoxins, glutaredoxins, and peroxiredoxins–molecular mechanisms and health significance: from cofactors to antioxidants to redox signaling. Antioxid. Redox Signal. 19, 1539–1605 (2013).
Он выполняет как защитные функции для защиты от физических и химических причин окислительного стресса, так и анаболические и восстановительные функции после травм.
Стационарные градиенты и управление ими.
Клеточный метаболизм характеризуется устойчивыми состояниями и переходами между ними в ответ на изменяющиеся условия. Важно отметить, что между клетками и субклеточными структурами существуют градиенты устойчивого состояния, а это означает, что в данном устойчивом состоянии в клетке для рассматриваемого агента существует пространственное распределение, напоминающее «ландшафт» с горячими точками, а не плоская концентрация по клеточное пространство.
Аналогично, химически активные вещества поддерживаются на устойчивом уровне, называемом «окислительно-восстановительным тоном» (который может быть определен для отдельных реактивных частиц, например, пероксидного и сульфидного тонов), благодаря жесткому контролю их источников и поглотителей. Для H2 O2 общая внутриклеточная концентрация оценивалась в диапазоне 1–10 нМ (reFs24,65), цитозольная концентрация оценивалась в 80 пМ (reF.66), концентрация в митохондриальном матриксе оценивалась в 5 –20 нМ (Х. Сайкс, неопубликованные результаты) и концентрация в просвет ЭР оценивался примерно в 700 нМ (reF.67).
Последняя из этих концентраций высока, поскольку ЭР является местом сворачивания белка с образованием дисульфидных мостиков, и на каждую образовавшуюся дисульфидную связь образуется одна молекула H2O2. Внеклеточная концентрация H2 O2 существенно выше и составляет около 1–5 мкМ в плазме крови68 (рис. 1). Таким образом, существует крутой градиент концентрации H2 O2 (в 100–500 раз) между внеклеточным и внутриклеточным пространством. Тиоредоксиновая система (см. также ниже) играет преобладающую роль в формировании внутриклеточные градиенты H2 O269. Контроль уровней H2 O2 в клетке также достигается за счет обмена между различными источниками, в основном включая контакты между ЭР, митохондриями и пероксисомами (см. ниже). Динамика метаболизма H2 O2 доступна для анализа с помощью генетически кодируемых флуоресцентных белковых индикаторных зондов.
Fransen, M. & Lismont, C. Redox signaling from and to peroxisomes: progress, challenges, and prospects. Antioxid. Redox Signal. 30, 95–112 (2019).
Lismont, C., Revenco, I. & Fransen, M. Peroxisomal hydrogen peroxide metabolism and signaling in health and disease. Int. J. Mol. Sci. 20, ijms20153673 (2019).
Appenzeller- Herzog, C. et al. Transit of H2 O2 across the endoplasmic reticulum membrane is not sluggish. Free Radic. Biol. Med. 94, 157–160 (2016).
Bestetti, S. et al. Human aquaporin-11 guarantees efficient transport of H2 O2 across the endoplasmic reticulum membrane. Redox Biol. 28, 101326 (2019).
Yoboue, E. D., Sitia, R. & Simmen, T. Redox crosstalk at endoplasmic reticulum (ER) membrane contact sites (MCS) uses toxic waste to deliver messages. Cell Death. Dis. 9, 331 (2018). 75. Dooley, C. T. et al. Imaging dynamic redox changes in mammalian cells with green fluorescent protein indicators. J. Biol. Chem. 279, 22284–22293 (2004).
Первым зондом, использующим H2 O2-чувствительный домен OxyR, был HyPer76 (см. reFs77–79 о текущем статусе в этой быстро развивающейся области. С помощью этих сенсоров и других инструментов теперь можно исследовать гетерогенность реакций отдельных клеток на окислители (например, в клеточном цикле).
Huang, B. K., Ali, S., Stein, K. T. & Sikes, H. D. Interpreting heterogeneity in response of cells expressing a fluorescent hydrogen peroxide biosensor. Biophys. J. 109, 2148–2158 (2015).
Будучи незаряженной молекулой, H2O2 способен проникать через биологические мембраны путем пассивной диффузии с определенной низкой скоростью. Однако было обнаружено, что H2O2 транспортируется с гораздо большей скоростью через водные каналы в мембране.
Henzler, T. & Steudle, E. Transport and metabolic degradation of hydrogen peroxide in Chara corallina: model calculations and measurements with the pressure probe suggest transport of H2 O2 across water channels. J. Exp. Bot. 51, 2053–2066 (2000). Discovery of H2 O2 transport across membranes by aquaporins.
Действительно, некоторые аквапорины (AQP3, AQP5, AQP8, AQP9 и AQP11) облегчают перемещение H2O2 через мембраны, поэтому их называют «пероксипоринами».
Bienert, G. P. & Chaumont, F. Aquaporin- facilitated transmembrane diffusion of hydrogen peroxide. Biochim. Biophys. Acta 1840, 1596–1604 (2014). Groundbreaking work on the role of some aquaporins as peroxiporins.
Таким образом, характер скорости переноса H2O2 через клеточные мембраны способствует установлению стационарных градиентов. Работа над AQP8 выявила механизм шлюзования, включающий персульфидацию цистеина (RSSH), открывая возможность контроля градиента H2O2 с помощью пероксипоринов окислительно-восстановительным способом.
Bestetti, S. et al. A persulfidation- based mechanism controls aquaporin-8 conductance. Sci. Adv. 4, eaar5770 (2018).
Medrano- Fernandez, I. et al. Stress regulates aquaporin-8 permeability to impact cell growth and
Поскольку митохондрии являются важными источниками H2 O2, вопрос о том, выделяют ли митохондрии в интактных клетках H2 O2 как таковой, является предметом текущих исследований. До сих пор не было прямых доказательств высвобождения H2O2 из митохондрий in situ в цитозоль. С использованием нового сверхчувствительного генетически кодируемого индикатора внутриклеточной диффузии H2 O2, HyPer7, H2 O2 из митохондриального матрикса в цитозоль можно было продемонстрировать только при ингибировании тиоредоксиновой системы в межмембранном пространстве, что подтверждает сомнения в физиологической роли прямого Высвобождение H2 O2 из митохондрий интактной клетки86. Существует вероятность того, что окислительный сигнал от ЦЭТ, генерируемый в виде H2O2, преобразуется в альтернативный окислительный сигнал для экспорта во экстрамитохондриальное пространство; это требует дальнейших исследований. Аналогично, было высказано предположение, что митохондриальные мембраны могут содержать аквапорины (в частности, AQP8)87, но это также кажется спорным. Важность поддержания клеточных градиентов H2 O2 можно проиллюстрировать на примере состояния редукционного стресса88; это может отражать недостаточность H2 O2 для поддержания устойчивого состояния окислительного эустресса. На системном уровне связь на большие расстояния между тканями и органами с помощью окислителей с низким содержанием, таких как H2O2, представляется маловероятной из-за повсеместного присутствия систем их удаления. Таким образом, короткоживущие оксиданты в основном физиологически используются для внутриклеточной передачи сигналов, а их продукция может быть связана с нереактивными веществами, такими как гормоны и цитокины, например, эпидермальный фактор роста (EGF)48, для межклеточной коммуникации. Тем не менее, стоит отметить, что гидропероксиды липидов могут транспортироваться между тканями, связанными с липопротеинами, а иммунные клетки с высокой скоростью производства H2O2, такие как гранулоциты, обладают высокой подвижностью, что может позволить использовать оксиданты в межклеточной коммуникации.
Мишени редокс-сигнализации
Передача редокс-сигналов влияет на функцию белка, приводя к изменениям в выходных сигналах, активности ферментов, транскрипции генов, а также целостности мембран и генома, и это лишь несколько примеров (рис. 3).
Выводы
- Как уже упоминалось, окислительно-восстановительная передача сигналов может также осуществляться посредством окислительной модификации РНК. В этом разделе подчеркивается плейотропность АФК в физиологической передаче сигналов. Что касается общего количества клеточных цистеинов, расчет показал, что около 10–20% тиолов из полных 214 000 тиолов в клеточном цистеиновом протеоме легко окисляются в аэробных условиях90. К ним относятся сайты регуляции ферментов, транспортеров, рецепторов и факторов транскрипции, а также сайты аллостерических и макромолекулярных взаимодействий.
- Различные белки, включая элементы цитоскелета, белки теплового шока, каркасные белки, такие как 1433 и многие рибонуклеопротеины, содержат цистеины, которые очень чувствительны к окислению, что указывает на существование функциональных белковых сетей, находящихся под окислительно-восстановительной регуляцией. Загадочные цистеины в белках могут стать доступными за счет изменений в структуре белка при стимуляции фактором роста, как показано для EGF.
- Другие мишени-окислители (кластеры железо-сера (Fe-s) или остатки тирозина; вставка 2) также обладают потенциальными сигнальными функциями и дополняют разнообразие механизмов окислительно-восстановительной передачи сигналов. Список целей редокс-сигнализации слишком обширен, чтобы его можно было охватить исчерпывающе (обзоры см. в reFs).
- В количественном тканеспецифичном ландшафте протеома, регулируемого редокс-регулированием, называемом набором данных Oximouse, сети окисления цистеина недавно стали доступны для детального изучения..
.