Безалкогольная жировая болезнь печени, которая варьируется от простого печеночного стеатоза без воспаления до безалкогольного стеатогепатита, фиброза/цирроза и даже гепатоциномы печени, в настоящее время признается наиболее распространенной в развитых странах. Последние данные показывают, что распространенность НАФЛР в мире составляет 25,24%, частота HCC среди пациентов НАФЛР - 0,044% в год, а общая смертность печени среди НАФЛР - 1,177% в год.

Недавние исследования показали, что "множественные совпадения" множественных факторов способствовали инициированию и прогрессированию НАФЛР. Эти факторы включают накопление липидов печени, инсулинорезистентность, воспалительную реакцию, окислительный стресс, митохондриальную дисфункцию, кишечную микробиоту и генетическое наследие. Накопление триглицерида в гепатоцитах является первым шагом в развитии NAFLD. Инсулинорезистентность является решающим фактором в NAFLD, который усугубляет дисбаланс липидного обмена. Воспалительная реакция и окислительный стресс рассматриваются как важные механизмы гепатоцеллюлярной травмы и прогрессии заболевания у пациентов с NAFLD. В лечении наиболее часто используемый витамин Е не облегчает фиброз печени. Терапевтическое воздействие инсулиновых сенсибилизаторов на НАФЛРД вызывает споры.
Материалы и методы
1. Животные и лечение
Гетерозиготные мыши на фоне C57Bl/6 J спаривались для получения потомства в центре эксперимента на животных/биобезопасности животных - Ⅲ лаборатории Уханьского университета с контролируемыми условиями среды, которая включала световые/темные циклы влажности 12-6 ч, температурой 22°C. Для идентификации гомозигот, гетерозигот и диких видов ПЦР проводили генотипирование семяпроводов. Мыши-гомозиготы и самцы дикого типа, которым было около 8 недель, были случайным образом разделены на четыре группы: 1. ВТСР: Группа мышей C57BL/6 J питается стандартно; 2. КОСД: Nr2e1 гомозиготы получают нормальную диету; 3. WT-HFD: Мыши C57BL/6 J питаются с высоким содержанием жиров; 4. KO-HFD: Nr2e1 гомозиготы получают диету с высоким содержанием жира. Оставшиеся ткани печени замораживали в жидком азоте, а затем хранили при температуре -80 °C.
2. Биохимические анализы
Образцы крови центрифугировались при 3500 об/мин и температуре 4 °C в течение 10 мин. Сывороточная аланиновая трансаминаза и аспартат-аминотрансфераза, активность общего холестерина, триглицерида, холестерина липопротеина низкой плотности и холестерина липопротеина высокой плотности были определены с помощью автоматического биохимического иммунного анализатора. Неэстерифицированные сывороточные свободные жирные кислоты и уровни гликогена в печени измерялись с помощью комплектов ИФА. Уровень инсулина в сыворотке крови на голодание измерялся с помощью комплекта ИФА.
3. Гистологический анализ
Кусочки печени были срезаны до 3-5 мм. Для доступа к липидным каплям криосекцию печени окрашивали раствором Oil Red O, а срезы парафина обрабатывали для окрашивания гематоксилина и эозина после депарафинизации и регидратации для наблюдения патологических изменений, включая распределение липидов. При периодическом окрашивании кислотой Шиффа микротомы печени парафина инкубировали в 0,5% периодический раствор кислоты в течение 5 мин при нормальной температуре. Затем секции были окрашены гематоксилином. В PAS было показано накопление гликогена в печени. Патологоанатом слепо проанализировал окрашивающие горки HE на наличие стеатоза, лобкового воспаления, вырождения воздушно-шарошечной системы оценки активности NAFLD.
4. Иммуногистохимия (IHC)
Кусочки парафина печени были обезжирены. Горячие ломтики для ремонта антигенов выдерживали в предварительно нагретом цитратном буфере при температуре 96-98 °C в течение 15 мин. 3% раствор пероксида водорода добавляли и выдерживали при комнатной температуре в течение 10 мин для деактивации активности эндогенных ферментов. Слои промывались фосфатным буферным солевым раствором. Круг был нарисован вокруг ткани полностью с помощью тканевой щетки. 10% козлиную сыворотку добавляли для герметизации неспецифических участков в течение 30 минут при комнатной температуре. Первичные анти-Nr2e1 и IL-1β антитела инкубировали в течение ночи при температуре 4°C. Вторичное анти-кроличье вторичное антитело пероксидазы хрена выдерживали в течение 30 минут при температуре 37 °C. Для получения цветного хромогена использовали раствор DBA. Гематоксилин был перекрашен ядром в синий цвет. Пленка была обезвожена и опечатана.
Общие характеристики мышей Nr2e1 (-/-) и создание моделей мышей с ожирением, индуцированных HFD
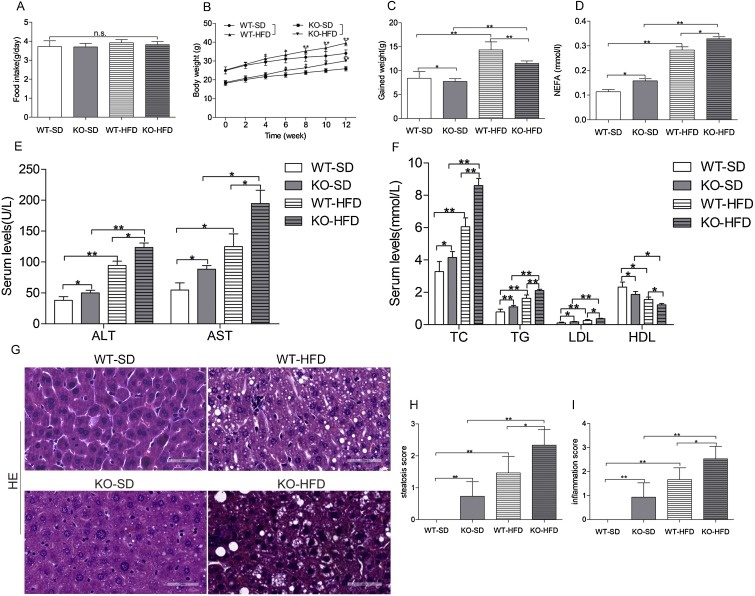
Было показано, что потеря функции Nr2e1 снижает массу тела мышей. По этой причине оценили характеристики влияния абляции Nr2e1 на регуляцию потребления пищи и массы тела. Nr2e1 KO у мышей наблюдалось немного меньшее потребление пищи, чем у мышей WT. Статистические различия между Nr2e1 (-/-) мышами и однотипными животными, питающимися одинаковым рационом, отсутствовали. По сравнению с мышами WT, мыши Nr2e1 KO набрали меньший вес на том же рационе.




















